
Freshwater Fishes of Iran
Species Accounts Revised:
06 May 2016
Back to Contents
Sparidae to Gobiidae
The sea breams or porgies are found in the Atlantic, Indian and Pacific oceans and comprise about 36 genera and about 130 species (Nelson, 2006; Eschmeyer and Fong, 2011). Some commonly enter estuaries and penetrate up rivers. Maximum length is about 1.2 m.
This family is characterised by a groove in the distal end of the premaxilla which accommodates the maxilla; the body is oblong to ovate and is compressed; the head is large with a steep upper profile; the preopercle margin is smooth; scales are weakly ctenoid, moderate in size and extend on to the cheeks and operculum; teeth are conical to incisiform and molar teeth are found in some at the rear of the jaw; there are no teeth on the vomer, palatines or tongue; the dorsal fin is continuous with an anterior spiny portion and a soft-rayed posterior portion about equal in size; with 10-13 spines and 10-15 soft rays respectively; spines fold into a groove; the anal fin has 3 spines (the second the largest) and 8-14 soft rays; branchiostegal rays 5-6; branchiostegal rays 5-6; and lateral line not continued onto the caudal fin but with enlarged scales near the head.
Many species in this family are hermaphrodites with male and female sex organs developing simultaneously, changing sex from male to female (protandry), or from female to male (protogyny). These fishes are often important as food or sought by anglers. Young fish may very different in colour to adults, usually being more vividly coloured with distinctive patterns. Most species are marine (see Marine List in Checklists in the Introduction) but a few enter fresh water and penetrate a considerable distance from the sea including one species in Iran.
Genus Acanthopagrus
Peters, 1855
Members of this genus have a compressed and moderately deep body, 4-6 enlarged incisiform teeth at the front of the jaws followed by 3-4 rows of molars, the second anal fin spine is longer than the third, there is a scaly sheath at the base of the dorsal and anal soft fins, and moderate-sized scales. Iwatsuki and Heemstra (2010) revised the genus in the western Indian Ocean.
Acanthopagrus latus
(Houttuyn, 1782)
Common names
شانك (= shanak), شانك زردباله (= shanak-e zardbaleh or yellowfin shanak).
[shanak, shaghoom, shaam, sha'm, shaem, sheim or sha-om in Arabic; yellow-finned porgy or seabream, yellow-finned black porgy, Japanese silver bream].
Systematics
Sparus latus was originally described from Japan.
Al-Hassan (1990) found differences in two meristic characters (pectoral and dorsal fin ray counts) but no differences in electrophoretic characters between populations from the Shatt al Arab and Khor al Zubair areas of southern Iraq. He concludes that there is only one stock of this species in southern Iraq as meristic variation may reflect environmental conditions.
Key characters
This species is the only sparid member recorded from Iranian freshwaters and is recognised by the dorsal fin spines alternately thick and thin and the colour pattern.
Morphology
Upper profile of head steep and convex back to above the posterior eye margin. The head bulges over the eye. Dorsal fin spines 11-13, soft rays 9-13. Anal fin with 3 spines, the second much stronger and wider than the third, and 8-9 soft rays. Pectoral fin branched rays 10-16. There is a strong spine in the pelvic fin and a well-developed axillary scale. Lateral line scales 41-46, or 48-50, or up to 55 depending probably on differing counting methods. The scales are vertical ovals with the anterior margin wavy where radii intersect. They have very fine circuli, moderate numbers of posterior radii, a subcentral posterior focus, and ctenii on the central part of the posterior margin extending inwards towards the focus. Four or five series of preopercular scales. The first pelvic fin ray is elongated as a small filament. There is a strong pelvic axillary scale. There are 3-4 scale rows sheathing the dorsal and anal fin bases. There are 4-6 compressed teeth in front of each jaw followed by 3-5 rows of molar teeth. The chromosome number is 2n=48 (Klinkhardt et al., 1995). The gut is an elongate s-shape.
Meristic values for Iranian specimens are:- dorsal fin spines 12, soft rays 10, anal fin spines 3, soft rays 8, pectoral fin branched rays 13, scales mostly lost.
Sexual dimorphism
This species is a protandrous hermaphrodite, being male early in its life and then becoming female later. Catches will include males, females and hermaphrodites, e.g. in Abu-Hakima's (1984a) study in Kuwait, there were 326 males, 343 females and 41 hermaphrodites.
Colour
Overall colour is a silvery-grey or silvery-white with the back darker and the belly yellowish. Scales each have a dark, brownish to golden spot at the base which line up to form apparent stripes along the flank. There is a dark blotch at the upper corner of the gill opening, on both the body and gill cover. There is a dark band over the head between the eyes and the edge of the operculum is dark. Dorsal fin spines are white and the membranes are grey, with dark margins between the spine tips. The soft dorsal fin is dark grey with a light orange tinge. There is a small back spot at the pectoral fin base and the fin is mostly hyaline with a light orange tinge. The anal and pelvic fins are a light yellowish-brown. The caudal fin is dark grey on the upper lobe and yellow on the lower with a black margin. The peritoneum is silvery brown in preserved fish with widely scattered melanophores.
Size
Reaches over 50 cm total length.
Distribution
Found from the Persian Gulf to Japan and north Australia. Recorded from the Helleh River in Bushehr Province, Iran (J. Holčík, pers comm., 1995), from the Bahmanshir River (Marammazi, 1995; Eskandary et al., 1999) and from Mahshahr creeks in the northwest Persian Gulf such as Jafari, Ghazaleh, Majidieh Petroshimi and Zangi, which are polluted (Savari et al., 2010, 2011; Movahedinia et al., 2011). Also from the Shatt al Arab in Iraq on the border with Khuzestan in Iran (Al-Hassan et al., 1989; Hussain et al., 1989).
Zoogeography
This marine species enters rivers in southern Iran and presumably freshwater stocks are maintained from this marine gene pool.
Habitat
The usual habitat is over sand and rock bottoms in the sea down to about 50 m, but young fish may enter estuaries and may penetrate considerable distances inland, although some fish remain at sea permanently. The frequency of penetration into Iranian rivers along the Persian Gulf coast is not known. Larger specimens are known to penetrate the Shatt al Arab in autumn, October to December, and this water body is an important nursery for this species, found there year round as young. Adults emigrate from January to March (Al-Hassan, 1990; Hussain et al., 1987). At a freshwater station on the Shatt al Basrah Canal with salinities up to 3.5‰, Al-Daham and Yousif (1990) found this species to be the second most dominant after Liza subviridis, comprising 7.1% by number and 10.9% by weight. Al-Daham et al. (1993) found young fish in the Shatt al Basrah mostly from April to October. Cage-cultured fish are reared at 14-31°C in Kuwait Bay (Abou-Seedo et al., 2003). Temperatures in Kuwait Bay during spawning are 14-18ºC during the winter months, a situation mirrored by Liza klunzingeri but not other species which begin to spawn in April (Abou-Seedo and Dadzie, 2004). Savari et al. (2010) showed that enzymatic indices could be used to evaluate ecological features and as biomarkers for pollution, Savari et al. (2011) showed that blood abnormalities and haematological parameters can be indicative of pollutants, and Movahedinia et al. (2011) showed that hormones could also be used in this regard.
Age and growth
A fast growing and hardy fish. Life in Iranian fresh waters has not been studied and information is about marine or estuarine populations.
A study of fish in the Shatt al Basrah canal, a man-made estuary of southern Iraq, was based on mostly small and immature fish (49-181 mm standard length) caught mostly in April-October. The length-weight relationship was W = 0.0511 L2.893, the dominant age group was 1+ fish, the maximum age was 3+ years, fish grew to 95, 155 and 215 mm total length in their first three years of life, and mortality (Z) was 2.23 (Al-Daham et al., 1993).
In an aquaculture experiment in Kuwait, fish more than doubled in weight over a 6 week period (Jafri et al., 1981). Males mature at a smaller size (12.3-14.2 cm) than females (24.3-26.2 cm) in cage-culture in Kuwait Bay (Abou-Seedo et al., 2003). However fish in the Shatt al Arab are usually less than 20 cm long and most are immature, in age groups 0 and 1. The length-weight relationship for both sexes was W = 0.0916 L2.6601. The lowest condition factors were found in April and May, possibly because fish were spent after spawning or were in lower condition after the winter (Hussain et al., 1987). In Kuwait, ages up to 14 years have been reported with ranges in von Bertalanffy growth parameters for the years 1981-1985 of L (cm) = 38.3-52.29, K = 0.169-0.298, and t0 -0.213- to -2.237. Total mortality values (Z) were 0.432-0.709, this range suggesting that, even for relatively large samples (92-314 fish), may be too small to provide reliable estimates of Z in species with a large overlap in age groups and where old fish are sampled only with difficulty. The length-weight relationship was total weight = 0.02874 x total length2.79198. Growth and mortality estimates based on all data were L∞ = 40.48 cm, K = 0.258, t0 = -0.965, tmax = 14 years and Z = 0.600 (Samuel and Mathews, 1987). Morgan (1985) gave values of L∞ = 43, K = 0.20 and Z = 0.97 but his study excluded fish over 30 cm and below 19 cm which accounts for the difference in mortality; and Samuel and Mathews (1987) had a value about 2.0 for Z when only fish 20-30 cm long were analysed.
Heydarnejad (2009) gave the length-weight relationship for an Iranian sample as W = 0.0451TL3.091.
Food
Freshwater food habits not known for Iran in detail but the one specimen examined contained plant fragments and scales of a cyprinid.
In a study of the recovering Hawr al Hammar, diet was 60% shrimps and 40% insects (Hussain et al., 2006). Feeds on echinoderms, worms, crustaceans, insects, bivalve molluscs and plants in the sea (Nasir, 2000; Al-Daham et al., 1993). Hussain et al. (1987) found crabs and bivalves to be the most important items by percentage in the Shatt al Arab Fish, shrimps and aquatic insects were also taken and there was significant seasonal variation, with shrimps and aquatic insects more important in December and spring. Hussain et al (1994) found bivalves in 91% of fish by number and weight in the Khawr az Zubayr. Al-Daham et al. (1993) found fish in the Shatt al Basrah fed on, in order of occurrence, crustaceans (decapods, amphipods, isopods, mysids, cladocerans and cyclopoids), fishes (Liza spp., Barbus (= Carasobarbus) luteus, Thryssa purava, eggs and scales), molluscs (Corbicula, Lymnaea, Tryonia and Sphaeriidae), algae (Oscillatoria, Syndera, Fragillaria and Cladophora), higher plants (Vallisneria, Ceratophyllum, seeds and roots) and aquatic insects (Corixidae, Hemiptera, Odonata and Coleoptera). Crustaceans were most important during July and November, molluscs in May and fishes during August. Hosseini (1998) examined food in coastal waters of Bushehr, Delvar and Rostami in the northern Persian Gulf of Iran and found 36.4% to contain crabs, 34.0% other fish and 13.4% shrimps. Snails and sea urchins were also eaten.
Reproduction
Al-Hassan et al. (1993) have shown for fish from the Shatt al Arab near Basrah, Iraq that haematological parameters vary with reproductive phase and between sexes. Cage-reared fish in Kuwait Bay have a prolonged spawning season from February to April. Fecundity there is up to 3,837,000 eggs. Spawning in the Shatt al Arab estuary is reported for April (Hussain and Ahmed, 1995) and Al-Daham et al. (1993) record a spawning season for the northwestern Arabian (= Persian) Gulf as January to April with a peak in February and March. Abu-Hakima (1984a) found the spawning period in Kuwait waters to be January to March with fecundity up to 2,152,993 eggs. This species is a protandrous hermaphrodite with males dominating in smaller size groups (22.3-24.2 cm) while females dominate in larger groups (24.3-26.2 cm) (Abou-Seedo et al., 2003). Samuel and Mathews (1987) give a spawning date of 1 December for their Kuwait sample. The gonadosomatic index was highest in February-March in Hosseini's (1998) study in coastal waters of the northern Persian Gulf.
The average absolute fecundity in coastal waters near Bushehr in Iran was 1,842,700, sex ratio was 1:1, and the spawning peak was January-February (Hossini and Savari, 2004).
Parasites and predators
None recorded.
Economic importance
A good food fish of high market value seen in bazaars along the Persian Gulf coast and in the Shatt al Arab. It was selling at U.S.$3.5-5.5 per kg in Kuwait about 1980, with 213 tons landed in 1995 for a value of U.S.$1,769,407 (Abou-Seedo et al., 2003). Experiments there indicate that this species can be farmed (Jafri et al., 1981; Abou-Seedo et al., 2003) and this has been proposed for Iranian waters in the Persian Gulf (Regunathan and Kitto, 2005). Experimental culture has been tried at Qeshm Island where a million larvae were produced in March with 100,000 larvae 2.0-3.5 cm long surviving in May (www.shilat.com, downloaded 7 June 2007). It is caught by trawls, handlines, in hadra (fixed stake nets) and gargoor (fish pots) and in the sport fishery in the Arabian (= Persian) Gulf (Samuel and Mathews, 1987; Carpenter et al., 1997).
Conservation
This marine species is fished commercially in the sea and populations there may be under some threat as a consequence. The status of freshwater populations is unclear as they appear quite rare and are presumably derived from marine populations at intervals.
Further work
The frequency of occurrence, detailed distribution and biology of this species in Iranian fresh waters needs study.
Sources
Iranian material: uncatalogued, 1, 62.9 mm standard length, Bushehr, about15 km above mouth in Helleh River (ca. 29º13'N, ca. 50º43'E).
Comparative material: BM(NH) 1974.2.22:1859, 1, 76.4 mm standard length, Iraq, Basrah (30º30'N, 47º47'E); BM(NH)1974.2.22:1858, 1, 76.3 mm standard length, Iraq, Beree (no other locality data).
Cichlids are found in fresh and brackish waters of Central and South America, Africa, Madagascar, the Levant, southern India, Sri Lanka and southern Iran. There are about 221 genera and about 1606 species (Nelson, 2006; Eschmeyer and Fong, 2011) but only 1 is found in Iran. Maximum length is about 80 cm.
Cichlids are distinguished by having only a single nostril on each side; practically all other fishes have two nostrils. The lateral line is in 2 parts, an anterior and higher portion ending under the soft dorsal fin and a lower, mid-flank, posterior part beginning below where the first part ends and continuing to the tail base, usually 7-25 spines in the dorsal fin followed by 5-30 soft rays, anal fin usually with 3 spines, but some species have 4-9 or 12-15 spines, followed by usually 4-15 soft rays, scales ctenoid or cycloid, extending onto the head, a specialised pharyngeal bone in the throat breaks up food by pressing it against a hard pad on the skull base, mouth dentition highly specialised in relation to diet with scraping (for algae on rocks), pointed (to seize fish), crushing (for hard-shelled molluscs), winkling (for removing snails from their shell or picking eyes) or reduced and embedded (for egg eating). Body form varies greatly between species and many are colourful and highly prized as aquarium fishes. They are often highly territorial, defending a breeding area against all invaders. They are most common in still waters where there are branches, rocks or plants. Some cichlids are important food fishes but they have attracted scientific attention for their elaborate breeding behaviour and evolutionary history. Certain cichlids, for example, are mouthbrooders, carrying eggs and fry in their mouths to protect them, while others spawn on the substrate, build nests or nourish young from a skin secretion. African lakes contain rich species flocks of cichlids which show various feeding behaviours. How these species arose and adapted to different ways of life have been important to scientists in understanding the mechanisms of evolution and adaptation.
Murray (2001) reviews the fossil record and the biogeography of the family and suggests an origin less than 65 MYA in the Early Tertiary in contrast to other studies that give an origin over 130 million years ago. Their salinity tolerance has enabled them to cross marine barriers.
The genus is monotypic so its description is subsumed under the Species Account below.
Iranocichla hormuzensis
Coad, 1982




Common names
mahi-e karoo, siklid Irani, siklid-e Hormuz, cichlid-e Hormuz.
[Hormuz cichlid, Iranian cichlid]
Systematics
Trewavas (1983) relates this species to Danakilia Thys van den Audenaerde, 1968, a genus with two species (Stiassny et al., 2010) from northeast Africa, on the basis of the low gill raker count, lower pharyngeal bone and teeth morphology, and morphometric characters such as a deep preorbital depth, long snout, head length and the small eye. Trewavas (1983) places Iranocichla in Danakilia but also agrees with Coad (1982a) that Iranocichla (and Danakilia) may be related to species in the genus Tristramella Trewavas, 1942 of the Jordan River basin and that this requires further investigation. Schwarzer et al. (2009) found molecular evidence for the relationship of Iranocichla and Tristramella (DNA material of Danakilia was lacking). I retain Iranocichla as a distinct genus until these relationships are examined more closely as some of the characters used to relate Danakilia and Iranocichla may be common responses to temperature and salinity extremes. In addition, Trewavas (1983) suggests a possible relationship of Danakilia with Oreochromis alcalicus (Hilgendorf, 1905) of the African Rift Valley. This species too is found in waters of high temperature and mineral content. Klett and Meyer (2002) group this genus with Sarotherodon, Oreochromis and Tristramella on mitochondrial NADH dehydrogenase subunit 2 gene sequences.
The type locality is the "Mehran River at 27°04'N, 54°35'E, Hormozdgan Province" (Coad, 1982a). The holotype, 94.2 mm standard length, is a female with eggs in the mouth held in the Canadian Museum of Nature, Ottawa under CMNFI 1979-0408A (see figure above). Paratypes are CMNFI 1979-0408B, 15, 24.3-86.5 mm standard length, same locality as the holotype and CMNFI 1979-0139, 35, 29.6-95.2 mm standard length, stream in Rasul River drainage between Chahar Berkeh and Tang-e Dalan, ca. 27°25.5'N, 54°59'E, Fars-Hormozgan border. Paratypes were deposited in the British Museum (Natural History), London under BM(NH) 1981.1.12:1-2 (2 specimens), Muséum national d'Histoire naturelle, Paris under MNHN 1981-107, 108 (2), California Academy of Sciences, San Francisco under CAS 47324 (2), the Royal Ontario Museum, Toronto under ROM 36389 (1) and the University of British Columbia, Vancouver under BC 81-1 (1).
Key characters
This is the only cichlid species in Iran, easily recognised by the single nostril opening on each side of the head.
Morphology
This cichlid is uniquely characterised by a nearly circular dental field on the lower pharyngeal bone, the teeth there being of uniform size and not enlarged medially and by cheek, operculum, belly, isthmus and area between the pectoral and pelvic fin bases naked or poorly scaled. Other significant characters are the posteriorly rounded dorsal and anal fins, short pectoral fins not reaching the vent, cycloid scales with granular posterior circuli bearing rounded or irregular protuberances, inferior apophyses for support of the swimbladder centred around the fourth vertebra (figured in Coad (1982a)), mesethmoid not meeting the vomer, modal vertebral count 29, median length of lower pharyngeal bone 31.8-40.9% (mean 35%) length of head, and pharyngeal blade/median length toothed area 0.6-1.0, mean 0.8.
Scales are regularly arranged on the flanks except that in some large specimens the regular scale rows are interspersed with irregularly distributed smaller scales, particularly on the upper flank. Scales may be absent entirely from the head, sparse above the lateral line anteriorly and on the belly posterior to the pelvic fins, absent from the dorsal and anal fin bases, absent from between the pectoral and pelvic fin bases and on the belly and isthmus anterior to the pelvic fins. However, in other specimens the head may be scaled dorsally to above the eyes, with scales variably imbricate, there may be 2-3 rows containing 4-7 minimally or non-imbricate scales on the cheek which is never completely scaled. The dorsal border of the opercle may have two large scales next to each other and a single scale may be present over the centre of the subopercular bone. Scales may be present on the whole belly, isthmus and between the pectoral and pelvic fin bases, but they are minute, embedded, and non-imbricate. Their extent and number varies between individuals. Small to minute scales, numbering up to about 20, are present on the caudal fin base, extending distally onto the fin membranes for more than half the fin ray length in some specimens.
Flank scales below the mid-point of the spiny dorsal fin and beneath the upper lateral line are cycloid or very weakly ctenoid. The focus is central and there are 9-14, mean 12.4, radii on the anterior field based on 5 scales from 7 adult specimens 59.2-87.1 mm standard length. Posterior circuli are granular so the exposed scale surface has rows of rounded or irregular protuberances.
The gut is a tightly coiled spiral with its apex ventral. Gut length in 5 specimens (59.2-90.5 mm standard length) is 6.8-8.3, mean 7.6, times the standard length. Gill rakers are short and rounded, reaching the adjacent raker or a little further when appressed.
The pharyngeal apophysis is of the Tilapia type (Greenwood, 1978). The mesethmoid does not meet the vomer, the intervening space being cartilaginous. Pores at the openings of the cephalic lateral line canals on the preorbital and preoperculum are single not multiple. The inferior apophyses for support of the anterior end of the swimbladder involve vertebrae 2 to 5, the fourth vertebra being involved in 8 out of 10 fish examined.
Teeth in the jaws are often irregularly arranged so that 4 rows are found in some places in both jaws. In some individual fish where teeth are regularly arranged there are 3 rows in the upper jaw and 4 rows in the lower jaw. Number of rows decreases laterally to one at the rictus. The outer row teeth are bicuspid with the lateral cusp the smaller, while inner row teeth are tricuspid, with the central cusp the most prominent. The upper jaw has more teeth than the lower jaw.
The diploid chromosome number is 2n=44, comprising 25 submetacentic, 18 subtelocentric and 1 metacentric chromosomes with an arm number of 70. The chromosome count may indicate a relationship to the Levantine Tristramella (Esmaeili et al., 2006).
Scales in upper lateral line 17(1), 18(1), 19(2), 20(8), 21(9), 22(10), 23(7), 24(4), 25(2), 26(1) or 29(1); scales in lower lateral line 9(6), 10(17), 11(14) or 12(9); total scales in lateral series 28(1), 29(2), 30(4), 31(9), 32(13), 33(5), 34(4), 35(4), 36(3) or 40(1); scales around caudal peduncle 16(15), 17(13), 18(15), 19(2) or 20(1); precaudal vertebrae 14(2), 15(53) or 16(11); caudal vertebrae 13(26), 14(35) or 15(5); total vertebrae 28(19), 29(40) or 30(7).
Dorsal fin spines 14(6), 15(46) or 16(14); dorsal fin branched rays 9(2), 10(36) or 11(28); anal fin branched rays 6(7), 7(20), 8(38) or 9(1); pectoral fin branched rays 11(42) or 12(24); and total gill rakers 14(6), 15(9), 16(24), 17(19), 18(6) or 19(1).
Sexual dimorphism
Head length is greater in females while pelvic fin length is smaller in females compared to males. Interorbital width is greater in males. Dorsal and anal fins are larger in males when expressed in terms of longest ray length in head length (Coad, 1982a). Colour differs as described below.
Colour
Live specimens are brightly coloured in spawning condition (based on aquarium photographs in Schulz (2004)). The male is brick-red on the lower sides and underside of the head with black on the dorsal head surface. The underside of the head may also be black. The belly anterior to the pelvics is black. The chin is white. The sides off the head have a few, scattered white spots but the body, dorsal and caudal fins are densely covered with white spots and blotches. Those on the dorsal fin are arranged in oblique rows and those on the caudal fin in bars. The anal fin has white spots also but these are not present distally. The pectoral fin has darkened rays but lacks spots. The pelvic fin has white spots proximally but less than the anal fin but is overall a dark black. Other reports and photographs (Svardal (2006) and Svardal and Svardal (2006)) show dominant spawning males to be black with brilliant turquoises blotches on the body but especially so on the fins. The female has an overall silvery colour with up to 9 faint to moderate flank bars. Fins are yellowish. The dorsal fin has a black tilapia-mark on the posterior dorsal fin.
Overall body colour outside the spawning season is a light lime green, with an iridescent tinge to the posterior edge of the operculum and on the back. The dorsal fin has light, lime-green, oblique bars, the last one or two black-edged and spot-like. The peritoneum is black.
Preserved specimens have the following pigmentation. Young fish have a distinct tilapia-mark, a spot on the rays of the soft dorsal fin typical of these cichlid fishes. The spot is black and is surrounded by a hyaline ring. Occasionally a second spot is found posterior to the first spot. The principal spot is often retained in adult fish. Young also have 7-11 bars along the flank which are also retained by adults but are then less distinct. In adults the dorsal fin rays and membranes are covered with melanophores interspersed with hyaline spots and irregular blotches. Wavy, oblique bars are found posteriorly on the soft dorsal fin in some specimens. The caudal fin has a series of about 7 narrow bars in some male specimens while females are uniformly grey. The anal fin is narrowly barred with up to 6 vertical to oblique bars in some specimens, in others uniformly pigmented grey proximally fading to hyaline distally. Pectoral and pelvic fins are not barred and are lightly pigmented, the pelvics being the darker. The head and body, including the belly, are more heavily pigmented to give an overall brown colour, lightest on the belly anterior to the pelvic fins in females. Scales are not pigmented on their free margins, which are pale.
Some specimens may be quite dark, particularly the back and fins and strikingly the lips.
Size
Attains 11.09 cm standard length or 12.95 cm total length (Esmaeili and Ebrahimi, 2006). Lamboj et al. (2006) give 13 cm, presumably total length.
Distribution


The cichlid is restricted to rivers draining to the Straits of Hormuz in southern Iran (Coad, 1982a; Abdoli, 2000). Svardal and Svardal (2006) also map this species at 27.770°N, 54.999°E, slightly to the north of samples mapped here. The distribution mapped by Stiassny in Keenleyside (1991) following Berra (1981) is too far north. The map in Berra (2001) is more accurate. Abdoli (2000) records this species from the lower Minab basin, lower Hasan Langi, middle to lower Kul, Gowdar and middle to lower Mehran rivers.
Specimens kindly sent to me by H. R. Esmaeili in 1997 are from the Dozdan River at 27°26'N, 57°10'E, an eastwards extension into the Minab River basin. The cichlid was not collected there in the 1970s. The new record may simply be filling in a collecting gap, a natural range extension or possibly the result of an introduction.
Bleher (2011a) found only 2 of the 22 sites recorded by Coad (1982) to still have water, although I have observed river stretches drying up and re-connecting and not always the same stretches.
Zoogeography
Trewavas (1983) suggests that the ancestor of this cichlid was distributed across the Arabian Peninsula in the Late Pliocene/early Pleistocene when this area was more humid. Desiccation in the Pleistocene and Recent Periods then led to the extinction of the ancestor. A Miocene-Oligocene fossil "Tilapia" was reported from Jisan in southwest Saudi Arabia where cichlids are not now native (Brown, 1970) and would help support a southern distribution of cichlids as the origin of Iranocichla. However Trewavas (1983) reports that this fossil cannot be identified as a cichlid. Micklich and Roscher (1990) and Lippitsch and Micklich (1998) also report three species of what are presumably cichlids from southwest Saudi Arabia in the Baid Formation of Oligocene age at Ad Darb, Tihamat Asir. They belong to the basal grade of cichlids and to two different clades within the African assemblage. Whybrow and Clements (1999) record unidentified Cichlidae from the Early Oligocene from the coastal trip of Dhofar, Sultanate of Oman with a date of 33 MYA. Murray (2001) reviews these and other cichlid fossil material and the identity of Omani material as cichlids appears questionable. Southwest Saudi Arabian material is more clearly cichlid but does not point to a continuous distribution eastwards across the Arabian Peninsula. However, Bănărescu (1992b) considers that a common ancestor to Iranocichla and the Levantine Tristramella evolved in the Arabian Peninsula from African forebears in the Miocene, the latter lineage extending its range northwards to the Levant and the former eastwards to Oman and southern Iran, the Straits of Hormuz not then being in existence. Murray (2001) gives an earliest date for colonisation of Iranocichla ancestors to be the Middle Miocene when southern Iran rose above sea level. She does not consider a coastwise dispersal through brackish waters of Arabia to be a possible route as cichlids are not found there today but indicates a route through the Tethys Sea/Indian Ocean could be possible.
Coad (1982a) suggests another hypothesis. Iranocichla could be a relict of a once wider distribution across the Tigris-Euphrates basin in a northern arc rather than directly across the Arabian Peninsula. The absence of cichlids from southern Arabia today warrant this alternative hypothesis. Warm streams have probably been continually present in southern Arabia and support a limited fish fauna today. There is no apparent reason why cichlids should have become extinct there. Murray (2001) points out that Iranocichla lives at 40-400 m above sea level and is limited by mountains north of the present distribution, and so it must have arrived before the mountains attained their current height, to support Coad's hypothesis.
The headwaters of the Tigris-Euphrates basin are narrowly separated from the Levant Rift Valley today and at times in the past may have had direct exchanges of faunas (Kosswig, 1965; 1973; Krupp, 1987). The modern absence of cichlids from the Tigris-Euphrates basin may be explained by low temperatures. The effects of low temperature on Iranocichla have not been investigated but fingerlings of Tilapia aurea, which occurs naturally in the southern Levant Rift Valley, begin to die at 11°C and cease all motion at temperatures below 10°C (Chervinski and Lahav, 1976). Most of Syria, northern Iraq and the northern Arabian Peninsula have temperatures below 10°C in winter (Beaumont et al., 1976). Spot temperatures from southern Iran at the head of the Persian Gulf are about 12-13°C in January compared to 18-20°C around the Straits of Hormuz where Iranocichla occurs. Additionally the death rate of Tilapia aurea in fresh water is twice that in salt water at low temperatures: it may be pertinent that Iranocichla is found mainly in saline streams. This hypothesis can only be confirmed by fossil discoveries.
Habitat
The streams in which this species lives are subject to desiccation with continuous flow breaking up into isolated pools. The survival of cichlid populations in these pools varies between years and some pools may be fishless in one year and populated in another.
The area around the Straits of Hormuz is rich in salt domes and consequently most surface streams are saline, up to 80 mS. Cichlids are found in these streams but also in the Sar Khun oasis which is fresh with a conductivity of 1.6 mS. Apparently they can be transported at 10 mS as this is less stressful. Stream waters are cloudy to clear and colourless. Water temperatures in winter (November to March) range from 15 to 33°C and would be considerably higher in summer when air temperatures reach 45°C with no riparian shade and low water levels. Lamboj et al. (2006) and Svardal (2006) give water temperatures of 33-40°C and conductivity of 45-75 mS.
Streams are 1 to 50 m wide and consist of alternating riffles and pools with occasional backwaters. The bottom is pebbles, sand or mud. Aquatic vegetation is restricted to encrusting algae.
Kiabi and Abdoli (2000) found this species to be, with Cyprinion watsoni, the commonest in Hormozgan Province.
Age and growth
Unknown. Esmaeili and Ebrahimi (2006) give a significant length-weight relationship based on 379 fish measuring 2.74-11.09 cm standard length. The a-value was 0.0349 and the b-value 3.047 (a b-value < 3 indicating a fish that becomes less rotund as length increases and a b-value >3 indicating a fish that becomes more rotund as length increases).
Food
Gut contents of 5 specimens (41.2-90.5 mm standard length) included only algae and diatoms suggesting food is scraped from rocks and from bottom deposits. This is consistent with an elongate gut and black peritoneum. Aquarium specimens eat algal tabs but also appreciate insects and fish remains.
Reproduction
This species is a mouth brooder. A breeding female and a male were caught in a backwater on March 18 of the Mehran River (the type series). This backwater was 1-5 m wide, maximum depth was 40 cm over a mud bottom, the water was cloudy and highly saline (40mS) and temperature ranged from 26°C at the mouth of the backwater to 33°C at its head. Eggs in fish taken in November and January are small so the breeding season is deduced to be around March. Five eggs ranged in length from 3.2 to 3.8 mm, mean 3.6 mm and in width from 2.4 to 2.7, mean 2.5 mm. Total number of eggs from 2 females, 59.0-59.2 mm standard length was 36 and 38 respectively. Eggs are yellow-orange in preserved fish.
A female 116.9 mm standard length from the Mehran River had 153 larvae in her mouth, ranging in length from 9.6 to 10.9 mm (H. R. Esmaeili, pers. comm., 6 October 2005; Esmaeili et al., 2009). Esmaeili et al. (2009) found a sex ratio biased towards males in May and June, presumably because males were defending nests and easily caught, or possibly differential survival of the sexes. They suggest that the breeding season begins in March and lasts until the end of June with a peak in May. Eggs attained 3.76 mm and fecundity reached 151 eggs with a relative fecundity of 5.4 eggs per gram body weight. Esmaeili et al. (2010) detail gonad morphology and histology and confirmed peak spawning in May.
Schulz (2004) observed fish in the field and found each male occupying a territory defending a nest about 1 m from each neighbouring nest. The nests were made on light grey, fine sand and consisted of a pit approximately 15 cm in diameter. The pit was black because of anoxic conditions below the sand surface. The actual nest was about the same as the body length of the fish (8-10 cm) and lay at the centre of the pit. The pit was surrounded by a rim about 1.5 cm high with an internally indented margin. Simpler pits are built where building materials are unavailable. Females were present in schools in deeper water in the river centre. Individual females swam purposefully to the nest defended by the male. The male directed the female to the nest centre with folded up fins while the female spread her fins and showed radiating colour changes. Spawning occurred immediately and neighbouring males intervened continuously at a speed that did not allow full analysis of the movements. A defending male would chase away an intruding male allowing another male into the unprotected nest to mate with the female. A clutch of eggs was always inseminated by a whole group of males.



Parasites and predators
Piscivorous birds have been observed along the streams where the cichlid is found. Ansary et al. (2010) record the anisakid nematodes Contracaecum sp. and Phocanema sp. from this fish in the Mehran River.
Economic importance
Saadati (1977) suggests that this salt-tolerant species could be a valuable resource if introduced into the saline and fishless waters of internal basins. However this is not advisable since the native fauna, evolved in a fishless environment, could be devastated before it has even been documented. Esmaeili et al. (2009) note that it is eaten by local people when available in large numbers in spring. It is now an aquarium fish in Germany (Schulz, 2002; 2004a; 2004b; Oliver Lucanus, pers. comm., 23 January 2004; Lamboj et al., 2006; Svardal, 2006; Svardal and Svardal, 2006) and juveniles sell for about $80 each (www.fishlore.com/fishforum/freshwater-beginners/67104-see-any-steals.html, 21 March 2010). Articles in aquarium magazines give photographs in spawning condition, including mouth-brooding, and details for their maintenance, including water with a conductivity of 50-70mS/cm NaCl or sea salt mixture, tank water changed once a week, vegetarian food tabs (containing the blue-green alga Spirulina), and a temperature of 20-35°C, optimally 27°C. It has been noted that males, in continually defending a nest and courting, "wear out" earlier than females (Thomas Schulz, in litt., 12 October 2006).
Conservation
Axelrod (1993) states that "as pollution chokes the waters of southern Iran, we can expect this fish to disappear very quickly" and "Because of the restricted range of this fish and the continual warfare and oil pollution in the area, Iranocichla hormuzensis may well be on its way to extinction - if it is not gone already". Bailey (2006) apparently repeats this. However its habitat is mostly saline streams which cannot readily be used for agriculture or industry. The surrounding area is not industrialised, nor likely to be, and was never a war zone so pollution is not a problem for this species.
Flash floods are probably a significant problem as water drains rapidly off vegetation barren land. The scouring action may well displace or strand cichlids. Mouth brooding offers protection against floods and against associated fishes.
Svardal (2006) and Svardal and Svardal (2006) give details of capture, transport and aquarium care of this species.
Further work
Further investigations into the biology of this species are needed.
Sources
Rabbaniha (1993a, 1993b, 1994) gives Farsi accounts of this species and cichlids in general. The account is based principally on Coad (1982a).
Type material: See above, CMNFI 1979-0408A, CMNFI 1979-0408B, CMNFI 1979-0139, BM(NH) 1981.1.12:1-2, MNHN 1981-107, 108, CAS 47324, ROM 36389 and BC 81-1.
Iranian material: CMNFI 1979-0138, 17, 25.6-97.3 mm standard length, Fars-Hormozgan border, stream in Rasul River drainage (ca. 27º32'N, ca. 54º58'30"E); CMNFI 1979-0140, 48, 24.2-59.4 mm standard length, Hormozgan, stream east of Kichal, Kul River drainage (27º14'N, 55º46'30"E); CMNFI 1979-0141, 5, 15.0-21.0 mm standard length, Hormozgan, Kul River (27º17'30"N, 56º03'30"E); CMNFI 1979-0142, 65, 21.3-82.8 mm standard length, Hormozgan, Baghu River (27º17'N, 56º28'E); CMNFI 1979-0143, 3, 25.0-26.7 mm standard length, Hormozgan, marsh in Hasan Langi River drainage (27º21'N, 56º50'30"E); CMNFI 1979-0148, 5, 33.2-92.7 mm standard length, Hormozgan, Sarzeh River (27º30'30"N, 56º15'30"E); CMNFI 1979-0181, Hormozgan, Kul River (27º17'30"N, 56º03'30"E); CMNFI 1979-0184, 12, 31.4-50.4 mm standard length, Hormozgan, effluent of Rasul River (27º11'N, 55º42'E); CMNFI 1979-0185, 7, 26.1-59.8 mm standard length, Hormozgan, stream in Rasul River drainage (27º06'N, 55º45'E); CMNFI 1979-0187, 2, 63.9-64.6 mm standard length, Hormozgan, stream at Sar Khun (27º23'30"N, 56º26'E); CMNFI 1979-0406, 5, 33.4-59.2 mm standard length, Hormozgan, stream north of Bandar-e Charak turnoff (26º48'N, 54º18'E);CMNFI 2007-0053, 17, 28.0-59.9 mm standard length, Hormozgan, Sarzeh River (ca. 27º36'N, 56º15'E); CMNFI 2007-0057, 14, 30.2-85.5 mm standard length, Hormozgan, Mehran River below Bastak (ca. 27º05'N, ca. 54º05'E); CMNFI 2007-0058, 2, 64.8-83.0 mm standard length, Fars, headwaters of Gowdar River (ca. 27º24'N, ca. 54º16'E)
Genus Oreochromis
Günther, 1889
Oreochromis niloticus
(Linnaeus, 1758)

Introduced to the Tigris River basin in Iraq but did not apparently survive winterkill (Herzog, 1969). Mutlak and Al-Faisal ( 2009), however, record O. aureus (possibly O. niloticus) from Basrah in southern Iraq and this species could easily become established in Iran. No Iranian record confirmed as yet. Red tilapias (Oreochromis sp.) have been studied in aquaponic systems in Iran so there is a potential for an exotic release (Rafiee and Saad, 2005). Fingerlings from Indonesia have been reared using saline waters at Bafgh, Yazd Province in 3 ton fibreglass tanks. Larvae were successfully grown to 2.0 kg at 28±1ºC (Iranian Fisheries Research Organization Newsletter, 57:2, 2009).
Tilapia zilli
(Gervais, 1848)

Introduced to the Tigris River basin in Iraq but did not apparently survive (Job, 1967). Redbelly tilapias are established in the Syrian Euphrates (R. Beck, pers. comm., 2000) and a recent report by Beshar Abd Al-Hussain Al-Saadi (in litt., 10 October 2006) of a cichlid at Al Musayyib on the Euphrates River in Iraq may well be this species. Mutlak and Al-Faisal ( 2009), record this species from Basrah in southern Iraq and these could spread to Iranian waters. No Iranian record as yet. The Farsi name is تيلاپيا (= tilapia).
The gobies are a world-wide family found mostly in warmer marine waters although some species enter fresh water and others live there permanently (see also Marine List in Checklists in the Introduction). The number of species is high and this may be the most speciose fish family in the world with about 248 genera and about 1630 species, perhaps more (Eschmeyer and Fong, 2011). A diversity of gobies occurs in the Caspian Sea basin. Not all Caspian gobies have valid Iranian records but most will probably be found there. Several gobies penetrate southern waters of Iran from the Persian Gulf and Sea of Oman and are described here. Others will probably be discovered when more detailed surveys are made.
Gobies are easily distinguished by their pelvic fins being united as an adhesive or sucking disk or cup. Body form and coloration are diverse. The pattern of head canals, canal pores and neuromasts is distinctive and used in identifying and relating species (except in "Neogobius" (Pinchuk, 1991)). However the neuromasts may be sunken in narrow furrows or pits and completely covered by epithelium so they do not preserve well and this can lead to confusion in identifications (Zambriborshch, 1968). There is usually a short spiny dorsal fin (2-8 flexible spines) separated from, but close to, a soft dorsal fin. The soft dorsal fin and anal fin are longer than the caudal peduncle. Scales may be cycloid, ctenoid or rarely absent. No obvious lateral line. There are 5 branchiostegal rays. Gill membranes are connected to the isthmus and gill openings are moderate to wide, or very restricted in the mudskippers. The head is usually blunt and the mouth is usually large. Teeth are usually small and conical in one to several rows in both jaws. Miller in Miller (2003) gives a suite of osteological characters defining the family.
Most gobies are quite small (5-10 cm) and they are often very abundant. Maximum size is about 50 cm. Some of the world's smallest vertebrates are gobies from the Indian Ocean, mature at 8 mm. Others, however, are large and form part of fisheries in both the Caspian Sea and the Indian Ocean. They are not significant food fishes in Iran. Gobies tend to rest on the bottom and move in sudden, characteristic dashes. The male goby guards a nest. Food is crustaceans, worms, molluscs and small fishes. Many gobies are important in the aquarium trade since they are beautifully coloured, small and tough.
They are known generally as gav mahi (= cow fish) or sag mahi (= dog fish) or contain the word gel (= mud) in Iran. A general review in Farsi of the Caspian gobies is given by Aslaanparviz (1991).
The males of some Caspian species become black during the spawning season, their fins elongate, head shape alters and some even become naked. Loss of tubercles in adult male gobies of the genus Benthophilus makes it possible to identify only juveniles and females. The males build nests and guard the eggs. Life span of certain Caspian species is said to be as short as one year, e.g. some species of Benthophilus, Knipowitschia, and Proterorhinus. Neogobius pallasi, N. melanostomus and Ponticola gorlap consume the invasive ctenophore, Mnemiopsis leidyi, as much as 10-15% of total biomass in some areas (Mamedov, 2006).
The Black and Caspian Sea basins contain an endemic Sarmatian fauna of gobies. There are two main clades, the gobiine-benthophilines (or transverse gobiids) and the pomatoschistines (or sand gobies), that have probably been distinct for at least 40 million years. Miller (2001) and Miller in Miller (2003) reviews the evolutionary history of these two clades and their anatomical differences based on head papillae and osteology. The transverse gobiids include Mesogobius, Neogobius, Proterorhinus, Chasar, Anatirostrum, Benthophiloides, Benthophilus and Caspiosoma while the sand gobies include Knipowitschia and Hyrcanogobius. The Sarmatian fauna was separated from the Atlantic-Mediterranean fauna with the isolation of the Paratethys during the late Miocene Messinian salinity crisis as the Mediterranean dried. Partial flooding of the Mediterranean from the Paratethys in the early Pliocene allowed Sarmatian gobies to spread westwards. Within the Ponto-Caspian basin, evolution of species flocks was favoured by basin sub-divisions and rejoinings. The benthophilines may be a monophyletic group from these events.
Ahnelt and Duchkowitsch (2004) give information on the neogobiine stock. About 12-13 million years ago in the Middle Miocene, the Ponto-Caspian endemic and ancestral neogobiine stock may have differentiated from an Atlantic-Mediterranean gobiine stock. At this time the Paratethys was a sea with reduced salinity and a high level of endemism. The Neogobius-Proterorhinus stock developed independently from the recent Gobius stock that invaded the Mediterranean basin after that sea was restored about 5 million years ago in the Late Miocene.
Neilson and Stepien (2009a) using mitochondrial and nuclear genes detail the evolution and biogeography of the subfamily Benthophilinae restricting Neogbius to caspius, melanostomus and pallasi with other known Iranian species formerly in Neogobius placed in Ponticola. The Benthophilinae includes Babka, Mesogobius, Neogobius, Ponticola and Proterorhinus as well as the tadpole gobies (Anatirsotrum, Benthophiloides, Benthophilus and Caspiosoma). The origin of the genera Babka, Benthophilus, Caspiosoma, Mesogobius, Neogobius, Ponticola and Proterorhinus ranges from 4.29 to 6.25 MYA and the paper gives divergence times for major lineages in relation to geological events in the Ponto-Caspian. These events include connections with, and isolation from, the World Ocean and salinity changes in a range of 1-30 p.p.t. over the last 5 million years. Most genera diversified about 5 MYA when the Black and Caspian seas separated. More recent separation events occurred during the Pleistocene glaciations Proterorhinus diverged from Mesogobius about 6.18 MYA, the Ponticola clade diverged about 4.51-4.86 MYA, and freshwater Proterorhinus emerged 1.18 MYA.
The principal recent works on the systematics of Caspian gobies are by V. I. Pinchuk, D. B. Ragimov, Ye. D. Vasil'yeva, H. Ahnelt and P. J. Miller. Earlier works are by B. S. Iljin (also spelled Il'in or Ilyin). Later molecular studies are cited above.
Other gobies in Iran are the familiar tropical mudskippers which
can move quickly over land, using the muscular-based paired fins to
row across mud, and some can even clasp and climb mangroves. They can live out
of water because the gill openings are small to prevent desiccation of
the gills, oxygen can be taken into the chamber and absorbed through
the gills and chamber wall, and they can also absorb oxygen through
their skin. They often rest with the tail immersed in water for this
purpose or roll around in shallow water to moisten themselves. They
may live entirely in water, or will come onto land even when there is
enough oxygen in the water. Their eyes are high on the head,
protruding and able to revolve independently, and have a movable lower
lid. The eyes are retracted periodically into small cups below the
head to moisten them. Such eyes are very effective as a means to watch
for potential enemies on land but their vision under water is blurred.
Mudskippers have elaborate reproductive behaviour which involves tail
standing, flip-flops, and fin displays. They are very territorial and
defend their territory against other mudskippers and crabs. They can
deliver a skin-breaking bite to humans even though they are only about 15 cm long!
This genus comprises only a single species and so its characters are those of
the species. The snout is very distinctive and details of neuromasts are not
given here as they are not needed in identification, although of importance in
relating the genus. The genus is closely related to the tadpole goby clade
comprising Benthophiloides-Caspiosoma-Benthophilus and details are give
in Miller in Miller (2004). This author also gives an alternative terminology
for the arrangement of neuromasts than that of Ahnelt et al. (2000).
Anatirostrum profundorum
(Berg, 1927)

Common names
gavmahi nuk-ordaki (= duckbill goby), gav
mahi-ye poozehderaz.[duckbill tadpole goby].
Systematics
This species was originally described in the genus Benthophilus Eichwald, 1831 by Berg (1927) but later Iljin (1930) erected a new genus because of its unusual and distinctive morphology. The type locality is the Caspian Sea at 37°58'N, 52°22'E at a depth of 294 m (but see below).
There are 14 fish under ZISP 23134 recognised as syntypes in Ahnelt et al. (2000) although Berg (1927) mentions 15 fish in his description. Ragimov (1985) states that Berg described this species from a single young specimen and also visually observed 15 others for a total of 16 in the type series.
Key characters
The duckbill tadpole goby is characterised by the elongate and flattened head which is similar to a duck's bill. Unlike gobies of the genus Benthophilus the body has minute platelets and granules, there are 8 rather than 6, vertical, suborbital series of pit organs, no postorbital occipital series, and no chin barbel or cheek flaps behind the jaw angle (Ahnelt et al., 2000).
Morphology
First dorsal fin with 3-4 spines, usually 4, second dorsal fin with 1 spine followed by 8-11, usually 10, soft rays. Anal fin with 1 spine followed by 8-11 soft rays. Pectoral fin rays 14-16. Gill rakers on the posterior part of the arch are very short and anteriorly are minute. Pit organs on the side of the head are papilliform and clearly visible with the naked eye. Further details of anatomy are given by Ahnelt et al. (2000).
Iranian specimens had the following meristics:- first dorsal fin with 4(4) spines; second dorsal fin with 1(4) spine followed by 10(4) soft rays; pectoral fin rays 14(1), 15(2) or 16(1); anal fin with 1(4) spine followed by 11(4) soft rays; and total vertebrae 29(4).
Sexual dimorphism
Females may retain dermal granules when mature.
Colour
Overall, colour is a light grey or pale fawn fading to a whitish grey on the belly. Various speckles and melanophores are found on the back and upper flank. The dorsal, caudal and pectoral fins have dark grey speckles. The head sides from the snout to the cheek are dark with transversal suborbital papillae series whitish giving the impression of narrow light stripes below the eye and on the cheek. The peritoneum is black or densely covered in fine speckles.
Size
Reaches 11.2 cm, or 13 cm total length (Jolodar and Abdoli, 2004). Females may be larger than males (mean total length 84 mm versus 77 mm).
Distribution
Found in the southern Caspian Sea including Iranian waters (Berg, 1927; Ahnelt et al., 2000; Abdoli and Naderi, 2009).
Zoogeography
Known only from the Caspian Sea and one of the endemic Sarmatian fauna (see Family Account).
Habitat
Found to a depth of 294 m on white silt bottoms according to Berg (1927) but the data in ZISP states 244 sazhems (= 446.5 m). Recent Iranian material is from 45-80 m, at 9.7-16.4°C at 50 m (Ahnelt et al., 2000) and Jolodar and Abdoli (2004) state it lives mainly at 50-100 m depths in the south Caspian Sea.
Age and growth
Unknown.
Food
Unknown but the duck bill may be an adaptation for feeding on silt bottoms (Ragimov, 1986).
Reproduction
Apparently mature eggs reach 1.9 mm in diameter in the Iranian specimen.
Parasites and predators
Unknown.
Economic importance
This species is too rare to be of any economic importance.
Conservation
Conservation requirements are unknown. Kiabi et al. (1999) consider this species to be data deficient in the south Caspian Sea basin according to IUCN criteria.
Further work
More specimens need to be caught to assess its distribution, numbers, variation and biology.
Sources
Type material: See above (ZISP 23134).
Iranian material: CMNFI 1999-0023, 4, 76.1-79.1 mm standard length, Gilan, Caspian Sea off Astara (38º00'N, 49º30'E to 38º20'N, 50º00'E).
Genus Babka
Babka gymnotrachelus Caspian Sea basin but no Iranian record although Kottelat and Freyhof (2007) map it from the Iranian shore. Gobius macropus De Filippi, 1863 described from Lake Palestrom near Poti, Georgia is
a synonym.
Babka
macrophthalma
Iljin, 1927
(Kessler, 1857)
(Kessler, 1877)
Reported from the south Caspian Sea by Naseka and Bogutskaya but no confirmed specimen from Iran.
Genus Benthophiloides
Beling and
Iljin, 1927
Benthophiloides brauneri
Beling and Iljin, 1927
Originally described from the lower Dnieper River between Kherson and Kakhovka and the south Bug River between Novaya Odessa and Nikolayev, Ukraine and also recorded from the Caspian Sea basin but no Iranian record. Ragimov (1998c) and Miller (2004) give recent descriptions. Pinchuk and Miller in Miller (2004) question the validity of the Caspian Sea record for this species.
Benthophiloides turcomanus
(Iljin, 1941)

Caspian Sea basin, described from off Chikishlar at 37°45.5'N, 53°47'E
and southwest of Ulsky Bank at 38°05'N, 52°34'E,
Turkmenistan, but no Iranian record. Known only from the two type
specimens, now lost (Reshetnikov et al., 1997).
Miller in Miler (2004) places this species in Benthophiloides; formerly
it was in the genus Asra Iljin, 1941. Miller in Miler (2004) gives a
general map that encompasses the southeastern Caspian Sea including Iranian
waters but the only records are not in Iran.
Genus Benthophilus Recheck Benthophilus illustrations in this account for source (correct species)
The tadpole gobies are found in the Black and Caspian seas where there are about 20 species, 16 endemic to the Caspian (Pinchuk and Miller in Miller, 2004; Boldyrev and Bogutskaya, 2007). The general
Farsi name for fishes in this genus is گاو ماهي (gav mahi) or سگ ماهي (sag mahi), not repeated under each species description.
Members of this genus are characterised by the broad and flattened head, dorsal muscles not extending to the eyes, no sensory canals or pores, first dorsal fin with 2-4 rays, well-separated form the
second dorsal fin, the caudal fin has only 2 rows of papillae, head and body scaleless but with spinulose bony granules, scutes and platelets except in adult males which are naked (and cannot therefore
be readily identified), anterior nostrils developed as small tubes overlying the upper lip, no swimbladder, a longitudinal dermal fold usually present on each side behind the mouth corner, and presence
of a chin barbel. There is no pelagic larval stage and eggs are large and oligoplasmatic. Ahnelt (2003) discuss the unique specialisations in the postcranial skeleton of benthophiline gobies (which also
includes Anatirostrum and Benthophiloides) including reductions in vertebral numbers and arrangement of pterygiophores. Pinchuk and Miller in Miller (2004) review other characters that show
relationships of this specialised group within a larger tadpole-goby clade, the broad and flattened head being the most obvious.
Pinchuk and Miller in Miller (2004) give details of the head neuromast organs that characterise this genus, namely 6 transverse infraorbital rows, 1-4 before and 5s and 6s
well above the level of the hyomandibular row b; row a is absent; row 5s is distant from row 4 which does not extend above the level of row b; rows 5i and
6i are arranged successively but row 5i is mostly below the level of row 6i separating the latter from the posterior end of row d; dorsal supraorbital rows o
very short and well-separated in the dorsal midline; and supraorbital series p present in interorbit as 3 pairs of short transverse rows.
These fishes are found in brackish waters with a salinity up to about 20‰, in deeper waters, estuaries and coastal waters.
They can dig themselves into bottom sediments and are usually found on mud, silt
or sand. Food is insect larvae, crustaceans and molluscs. Life span of these tadpole gobies is only a year, some individuals maturing
at 6-7 months, called ephemery. Fish die after spawning, females earlier than males by 3-4 weeks (Boldyrev and Bogutskaya, 2004; 2007).
Eggs are laid inside an empty mollusc shell. Pinchuk and Miller in Miller (2004) and Boldyrev and Bogutskaya (2004; 2007) give descriptions and keys for all Caspian Sea species. The 5 species cited here are the ones with records from Iran although
it is likely other species will eventually be found along the southern Caspian Sea coast.
Benthophilus abdurahmanovi Endemic to the Caspian Sea, found in the North Caspian and reported from the
south Caspian Sea by Naseka and Bogutskaya (2009). No Iranian record.
Benthophilus baeri
Common names
gavmahi-ye Baer, gavmahi-ye tokmehsar.
[Ber comcaxulu in Azerbaijan; Baer pugolovka or Baer tadpole goby in Russian].
Systematics
Benthophilus Baeri was described from the Caspian Sea at Mangyshlak Peninsula, Kazakhstan and in the South Caspian.
A specimen listed as a syntype is in the Natural History Museum, London under BM(NH) 1897.7.5:16 (32.5 mm standard length) from St. Petersburg University (sic, ZISP?) with others in the
Zoological Institute, St. Petersburg (ZISP 2239, lectotype and ZISP 53665 (ex 2239) (4)) and SPU 395 (463/410) (1) (Eschmeyer et al., 1996; Pinchuk et al., 2004).
Lectotype: ZISP 2239. Paralectotypes: (11) BMNH 1897.7.5.16 [ex St. Petersberg Univ.] (1), SPU 395 [463/410] (1), ZIN 53665 [ex 2239] (4).
Key characters
This species has very large and high, conical tubercles arranged in rows, not all tubercles are spinous, dorsal row tubercles usually 11-17 (lower than other Benthophilus), ventral row tubercles
9-14 (lower than other Benthophilus), granules are mostly absent between body tubercles rows, interorbit with a shallow median groove, a chin barbel is present, there are 1-2 dermal barbels behind
jaw angle, the first dorsal fin has 1-2 spines, and dark bands are absent.
Morphology
First dorsal fin with 0-2 spines, usually 1 (Ragimov (1985) and Boldyrev and Bogutskaya (2007) but 2 according to Pinchuk et al. in Miller (2004)), second dorsal fin with 1 spine followed by
6-9, usually 7, soft rays. Anal fin with 1 spine followed by 5-9, usually 6-7, soft rays. Dorsal row tubercles 11-17, usually 13-15, lateral row 1-12, usually 7-9 (but sometimes completely absent), and
abdominal row 9-14, usually 11-12. Total vertebrae 24-27. Tubercles on the head and flanks are large and can bear 1-4 sharp spines but only some tubercles retain spines. The dorsal tubercle row
continues onto the head ending near the eyes while the lateral row is short, extending from a level between the dorsal fins and ending at the rear of the second dorsal fin. There may be a few small
granules scattered between the major row tubercles and on the head. There is a long chin barbel, about equal to eye diameter. There is one neuromast in row 7 behind the eye.
A single Iranian specimen had ?
Sexual dimorphism
The second dorsal and anal fins of large males are enlarged.
Colour
The back, upper flank and upper head are pale grey to pale brown with dark grey speckles, and often minute dark dots form dark areas. Some bands may be weakly expressed on the back and flank, more
obvious in immature specimens. The lower body and belly are whitish. The pectoral, second dorsal and caudal fins have dark grey speckles and bars while the other fins are pale.
Size
Reaches 8.6 cm for males and 6.9 cm for females.
Distribution
Found in the central and southern Caspian Sea, rarely in the northwest Caspian. Reported from Gorgan Bay, the southeast Caspian Sea, southwest Caspian Sea and south-central Caspian Sea in
Iran (Kiabi et al., 1999; map in Reshetnikov, 2002; Pinchuk et al. in Miller, 2004;
Abdoli and Naderi, 2009).
Zoogeography
A Caspian Sea endemic, part of a unique fauna poorly studied and collected in Iranian waters, as are all its relatives below.
Habitat
Found in depths of 5 to 81 m in littoral areas and also known to enter rivers in Russia (Reshetnikov et al., 1997). Found at 0.5-30.0 m in warm seasons and migrates to deeper water at 100 m in
winter (Boldyrev and Bogutskaya ,2007). During the spawning season they are absent from shallow waters (5-10 m) and deeper waters (30-50 m)(Pinchuk et al. in Miller, 2004).
Age and growth
Up to 4 age groups are known (Reshetnikov, 2002) although Pinchuk et al. in Miller (2004) indicate life-span may be only 18 months.
Food
Corophiid crustaceans and some molluscs such as gastropods are principal diet items. Cumaceans, amphipods and Nereis worms are also important food items (Pinchuk et al. in Miller, 2004).
Reproduction
Mature males measure 8 cm while a mature female from the same collection was 6 cm (Berg, 1948-1949) while Pinchuk et al. in Miller (2004) state that both sexes are mature by 4.0 cm, in the first
year of life. Eggs have a maximum diameter of 1.2-1.8 mm and a minimum diameter of 0.3-0.5 mm and fish up to 5.2 cm long produce up to 1860 eggs (Reshetnikov, 2002). Spawning takes place from the second
half of May to August and rarely into September with a second batch of eggs laid in July (Pinchuk et al. in Miller, 2004). Eggs are laid on silt and silt-sand bottoms, probably using shells, at
10-25 m (Pinchuk et al. in Miller, 2004).
Parasites and predators
None recorded from Iran.
Economic importance
This species is food for sturgeon and Sander species (Pinchuk et al. in Miller, 2004).
Conservation
Kiabi et al. (1999) consider this species to be data deficient in the south Caspian Sea basin according to IUCN criteria. Criteria include possibly few in numbers, limited range (less than 25%
of water bodies), absent in other water bodies in Iran, and absent outside the Caspian Sea basin.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
Type material: See above (BM(NH) 1897.7.5:16).
Iranian material:
Benthophilus casachicus Endemic to the Caspian Sea and found on the eastern Caspian coast, the Volga delta area,
and from
the south Caspian Sea by Naseka and Bogutskaya (2009). No Iranian record.
Benthophilus ctenolepidus
Common names
gavmahi-ye shafaf or shaffaf (= transparent or clear goby); gavmahi-ye Lankaran or gavmahi-ye Lenkoran.
[prozrachnaya pugolovka or transparent tadpole goby, shipogolovaya pugolovka or spiny-headed (?) tadpole goby in Russian; spiny-scaled tadpole goby; Lenkoranskaya pugolovka or Lenkoran tadpole goby in
Russian for the subspecies in the southern Caspian Sea].
Systematics
A subspecies reported from off the Iranian coast was Benthophilus ctenolepidus pinchuki Ragimov, 1982 but this is now recognised as a distinct species by Pinchuk and Miller in Miller (2004)
(see below). The lectotype of Benthophilus ctenolepidus as designated by Ragimov (1982) is in the Zoological Institute, St. Petersburg under ZISP 10897 with 3 paralectotypes lost and 1
paralectotype from ZISP in the Natural History Museum, London under BM(NH) 1897.7.5:13 (66.9 mm standard length) (Eschmeyer et al., 1996). The type locality is "yuzhnoi i srednei chastyakh'
Kaspiiskago morya" (= southern and middle Caspian Sea), or Caspian Sea, 40°08'N, 0°26'E off Baku, Azerbaijan (in Eschmeyer's "Catalog of Fishes" online, downloaded 22 August
2007). Records of B. magistri Iljin, 1927 in the Caspian Sea, as B. m. lencoranicus Ragimov, 1982, are this species (Boldyrev and Bogutskaya, 2007). The taxon Benthophilus magistri
lencoranicus Ragimov, 1982 was described from "raione protib Zelenogo bugra (38°10' s. sh.)" (= area opposite Green Hill, Turkmenistan). The holotype of lencoranicus
is in the Caspian Biological Station, Institute of Zoology, Azerbaijan under CBSIZA 546 (no. 2), paratypes under 546 (20) and additional non-type material in the Zoological Institute, St. Petersburg
under ZISP 23131 and 41912 (9) (Eschmeyer et al., 1996). Boldyrev and Bogutskaya (2007) were unable to locate IZA 546.
Key characters
The narrow head has a trough-shaped depression or groove on the occiput, the large tubercles are vertically elongate, curved and crest-like and have their rear edges fringed with a comb of spines,
dorsal row tubercles usually 30-33, abdominal row tubercles usually 24-26, temporal and occipital region without large tubercles but having granules, the body has tiny granules anteriorly interspersed
among the large tubercles, dermal filaments present or absent, a small but well-developed chin barbel is present, the dermal process behind jaw angle is lobed, narrow, with an acute protuberance, first
dorsal fin spines 3-4 (rarely 2), and back without brown bands.
Morphology
First dorsal fin with 3-4 spines, second dorsal fin with 1 spine followed by 8-11, usually 9-10, soft rays. Anal fin with 1 spine followed by 7-11, usually 8, soft rays. Total vertebrae 29-230. Dorsal
row tubercles 27-33, usually 30-33 (?) or 22-24 (Boldyrev and Bogutskaya, 2007), lateral row 11-22, usually 17-19, abdominal 20-26, usually 24-26 (22-24 in Boldyrev and Bogutskaya (2007)). Tubercles on
the head (when present) and flanks are large and bear sharp spines, the flank tubercles being angular in shape. The dorsal row of tubercles is incomplete, usually beginning below the first dorsal fin.
The first 3-4, and as many as 7, dorsal row tubercles are markedly smaller than the succeeding ones. The dermal plate or fold behind the mouth corner is large (1.2 times eye diameter),
oval, and without a wavy margin.
Sexual dimorphism
Undocumented.
Colour
Mostly transparent as the name suggests, a light to ash grey. The body bears dark lines and speckles but no strong dark bands, spots or blotches. The belly is yellowish-white. The dorsal and pectoral
fins have thin banding and the pelvic and and anal fins are pale.
Size
Reaches 10.7 cm.
Distribution
Found in the central, southwestern and southeastern Caspian Sea and in adjacent Iranian waters (Ragimov, 1965;
Abdoli and Naderi, 2009). Records are from Astara on the Azerbaijan-Iran border to Hasan Kuli in Turkmenistan and
adjacent waters.
Zoogeography
A Caspian Sea endemic.
Habitat
Found in depths of 0.5-74 m in littoral areas but deeper than B. magistri and B. macrocephalus. It feeds and spawns in 0.5-10.0 m and migrates to deeper water in winter. Occurs in higher
salinity areas at 12.4-13.0‰. Jolodar and Abdoli (2004) place it at 30-200 m depths in the south Caspian Sea basin. The pelvic fin is less developed, shorter and not as dilated than in these shallow
water species.
Age and growth
Life span is probably not more than 18 months with maturity at one year (Pinchuk and Miller in Miller, 2004).
Food
Food in northern Caspian specimens was found to include mostly corophiid amphipods, with some gammarids, other crustaceans, and gastropods (Pinchuk and Miller in Miller, 2004).
Reproduction
Spawning is thought to occur from spring to autumn and into winter (Pinchuk and Miller in Miller, 2004).
Parasites and predators
None reported from Iran.
Economic importance
None.
Conservation
Kiabi et al. (1999) consider this species to be data deficient in the south Caspian Sea basin according to IUCN criteria. Criteria include possibly few in numbers, limited range (less than 25%
of water bodies), absent in other water bodies in Iran, and absent outside the Caspian Sea basin.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
Based on Pinchuk and Miller in Miller (2004).
Type material: See above under B. ctenolepidus (BM(NH) 1897.7.5:13).
Benthophilus granulosus Endemic to the Caspian Sea and found in coastal areas of the North and eastern and western Middle Caspian. No Iranian record although Kottelat and Freyhof (2007) map this species from the Iranian shore.
Benthophilus grimmi A Caspian Sea endemic found in the western part of the Middle Caspian and the northern part of the South Caspian south to about 39º40'N,
and from the
south Caspian Sea by Naseka and Bogutskaya (2009). No Iranian record.
Benthophilus kessleri A Caspian Sea endemic found in coastal waters of the eastern Middle Caspian,
and from the
south Caspian Sea by Naseka and Bogutskaya (2009). No Iranian record.
Benthophilus leobergius
Common names
gavmahi-ye akhtari (= stellate or starry goby), gav mahi bacheh qurbaqei, gavmahi-ye bacheghoorbagheie, sebele.
[ulduzlu comcaxul or khazar julduzlu chomchakhulu in Azerbaijan; zvezdchataya pugolovka or stellate tadpole goby in Russian; Caspian starry goby, Caspian stellate goby].
Systematics
This taxon was originally described as a subspecies of Benthophilus stellatus (Sauvage, 1874). B. stellatus is now restricted to fish from the Black Sea. Note that views do conflict on
the status of this taxon and more studies remain to be done (Pinchuk et al. in Miller, 2004).
The subspecies leobergius (sic; leobergi has been used, see below) was first proposed by
Iljin (1949) and was thought not be available as no distinguishing features were given in
the description (Eschmeyer et al., 1996) although Berg (1948-1949) gives a description referring to "higher development of spiny tubercles" and colouration "less bright"
than then Black Sea form in his volume III dated 1949 (not 1948 as given in Eschmeyer et al. (1996)). Eschmeyer et al. (1996) consider that Berg may be the author of this taxon and Pinchuk
et al. in Miller (2004) concur. The holotype is probably in the Zoological Institute, St. Petersburg under ZISP 10891 (Eschmeyer et al., 1996). Berg (1948-1949) notes that the taxon is found
"in the freshened regions throughout the Caspian Sea, south to Astrabad Bay (No. 10891, 5 August 1874, 62 mm)". This may be the type locality, i.e. Gorgan (= Astrabad) Bay,
Iran. However this specimen may be merely recording the southernmost distribution of the taxon and a larger fish at 103 mm under ZISP 23128 labeled as from the Caspian Sea could be the holotype as it is
depicted in two views (Figure 858, p. 1115 in the Russian text and p. 196 in the English text, and Figure 859, p. 1116 Russian and p. 197 English). Both these fish may be syntypes.
Iljin (1949) simply
referred to "Kaspii", i.e. the Caspian Sea, as the type locality. Boldyrev and Bogutskaya (2007) selected ZISP 23128 as the lectotype. Benthophilus macrocephalus variety b (large-scaled)
Kessler, 1877 is a synonym of B. s. leobergius according to Berg (1948-1949). Benthophilus aculeatus Baer in Lukina, 1984 is possibly a synonym, with no types known (Boldyrev and Bogutskaya, 2007).
Kottelat (1997) considers, tentatively, that Benthophilus stellatus is restricted to the Black Sea basin. Furthermore, Kottelat points out that perhaps the Caspian Sea species is Benthophilus
leobergi Ragimov, 1978, leobergi since the species is founded on a personal name and should be emended to this spelling and Ragimov, 1978 since the name is a nomen nudum in Iljin (1949) (no
description or diagnostic characters (but see above)) until Ragimov (1978) makes the name available by listing characters distinguishing it from B. s. casachicus. Benthophilus casachicus
Ragimov, 1978 is then another Caspian Sea species.
Key characters
Distinguished by stellate tubercles, mostly spinous at the tip, tubercles rows on body well-defined but scattered on head, dorsal tubercles 25-30, abdominal tubercles 21-25, granules present
predorsally but mostly absent from head and body, interorbit without a median groove, chin barbel present but small (eye diameter or less), dermal process behind jaw angle obvious, 3-4 first dorsal fin
spines, and first dorsal fin patterned. and dark bands present.
Morphology
The snout is very long, 30-33% of head length, and the upper jaw projects. The head is wider than long. There are 3-4 neuromasts in row 7 (also in B. macrocephalus, usually 1 in other species).
First dorsal fin spines 2-6, usually 3-4, second dorsal fin with 1 spine and 6-11 soft rays, usually 8-10, anal fin with 1 spine and 7-9 soft rays. Pectoral fin rays 15-18. The gill arch is without
evident rakers and is irregularly tuberculate. Vertebrae number 28-31. There are 10-11, often 11, precaudal vertebrae (commonly 9 in other Benthophilus). There is a flap behind the jaw angle. The
gut is moderately long with several convolutions. The body bears 3 rows of very spinulose tubercles; the upper row numbering 25-30, usually 26-28 or 27-29 (authors vary), the lower 21-25, usually 23-25
or 22-24 (authors vary), and the dorso-lateral row 15-24, usually 18-20. The ventro-lateral row is usually absent.There are numerous small tubercles between the main rows on the flanks, particularly the
dorsal and lateral rows. The head surface has numerous small and large tubercles, arranged irregularly. Small tubercles or granules are even found on the medial half of the eyeball. There are 2
well-developed tubercles on the midline between the eyes (in 80% of specimens) as opposed to one or none in other Benthophilus.
The chromosome number of B. leobergi is 2n=44 with 46-48 chromosomal arms (Grigoryan and Vasil'ev, 1993; Vasil'yev and Grigoryan, 1993) or 2n=46 (Klinkhardt et al., 1995).
Iranian specimens had the following meristics: first dorsal fin spines 3(1) or 5(1), second dorsal fin with 1 spine and 9 soft rays (2), anal fin with 1 spine and 8 soft rays (2), and pectoral fin with
17(2) branched rays. The extent of tubercle rows is difficult to distinguish since their size gradually decreases posteriorly and their arrangement is irregular anteriorly. Dorsal row tubercles 31(2),
lateral row tubercles ca. 19(2), and lower row tubercles 23(1) or 24(1).
Sexual dimorphism
The pectoral fin is longer than the pelvic fins in males when spawning. Females have a very small chin barbel.
Colour
Overall colour is brownish to whitish-grey. The head surface and upper flank have large, dark brown spots and small dark speckles. The body has about 5 transverse bands which circle the first dorsal
fin, rear of the second dorsal fin and caudal fin base when viewed from above. Rows of speckles are found on the second dorsal, pectoral and caudal fins.
Size
Reaches 10.7 cm.
Distribution
Found in the Caspian Sea. In Iran, it is reported from Gorgan Bay (Abdoli, 1994a). the Safid River (Abbasi et al., 1999), the southeast Caspian Sea, southwest Caspian Sea and south-central
Caspian Sea (Kiabi et al., 1999;
Abdoli and Naderi, 2009). Jolodar and Abdoli (2004) consider it to be widely distributed along the Iranian coast but more abundant in Gomishan Wetland and Gorgan Bay. It is also recorded
between Kultuk and Astara in Azerbaijan, near the Iranian border (Ragimov, 1965).
Zoogeography
Endemic to the Caspian Sea.
Habitat
This species is found in the Caspian Sea and in brackish and fresh waters, and is widespread in coastal waters over sand and shell debris bottoms down to at least 100 m. Said to prefer salinities
below 9‰. Feeding and spawning occur in shallow water at 0.5-10.0 m, retreating to deeper water in winter (Boldyrev and Bogutskaya, 2007). It is said not to enter fresh waters unlike its relative
B. stellatus but is known from fresh water in the Volga delta. In spring and autumn it can be found in coastal zone at 1-20 m. It prefers warmer waters at 18-26ºC (Caspian Sea Biodiversity
Database, www.caspianenvironment.org).
e Age and growth
Young fish reach 1.83 cm in January and early February after spawning in autumn and reach their maximum size in July-August of the following year at 8.5-9.4 cm and 21.6-24.7 g. The spring spawners
reach 1.9-2.0 cm and 0.17-0.24 g in May-June and 3.5-5.9 cm and 1.1-5.4 g in July-August. With two spawning seasons, there are always fish of markedly different size and maturation stage
in any population. Life span is about 18 months (Ragimov, 1985a). Abdoli et
al. (2006) examined fish from the Gomishan and Miankaleh wetlands and found
a length (L)-weight (W) relationship of W = 0.0499L2.5 for males. Food
Diet is dominated by molluscs (bivalves and gastropods) but includes crustaceans, worms, insect larvae and small fishes. An Iranian specimen contained the remains of a crab.
Reproduction
In the southern Caspian spawning occurs at depths of 30-35 m and in October-December and March-April (in the northern Caspian it is in shallow water (1-5 m) and during summer months). Batch spawning
occurs and eggs number up to 2136 large and 1284 small with diameters 1.5-2.1 mm and 0.3-0.7 respectively (Ragimov, 1985a). Fecundity may reach 3500 eggs and maximum diameter 4.5 mm (Reshetnikov, 2002).
Males protect the nests; unprotected nests have their eggs eaten by crustaceans within 2-3 hours (Caspian Sea Biodiversity Database, www.caspianenvironment.org).
Parasites and predators
Sattari et al. (2004) record the nematode Anisakis sp. from
this species in Gilan. This fish is eaten by other fishes and by seals.
Economic importance
None.
Conservation
Kiabi et al. (1999) consider this species to be of least concern in the south Caspian Sea basin according to IUCN criteria. Criteria include medium numbers, medium range (25-75% of water bodies),
absent in other water bodies in Iran, and present outside the Caspian Sea basin.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
Iranian material: CMNFI 1971-0326A, mm standard length, ():
CMNFI 1970-0543, ();
3, 90.4-90.9 mm standard length, Mazandaran, Gorgan Bay and the Caspian Sea.
Benthophilus leptocephalus A Caspian Sea, deep-water endemic found in the western Middle Caspian south to 39º41'N and the eastern South Caspian south to Hasan Kuli in Turkmenistan. No Iranian record.
Benthophilus leptorhynchus A Caspian Sea, deep-water endemic known from the western coast of the Middle Caspian and the north of the South Caspian south to Baku. No Iranian record.
Benthophilus macrocephalus
Common names
gavmahi-ye vazaghi (= tadpole goby).
[iribas comcaxul in Azerbaijan; Kaspiiskaya pugolovka or Caspian tadpole goby in Russian; bighead goby].
Systematics
Eschmeyer et al. (1996) gave the publication date as 1788 (later 1787), Berg (1948-1948) and Pinchuk and Miller in Miller (2004) as 1787. The original description is dated 15. Mars. 1787 but
was probably published in the following year (not verified). The type locality is the Caspian Sea near the mouths of rivers and streams, and in small inlets, although the type of Gobius
macrocephalus was probably from the Volga delta. No type specimens are known.
Key characters
Distinguished by having moderate, stellate tubercles, spinous at their apex with usually 22-23 (but see below) in the dorsal row and 19-24 in the abdominal row, numerous granules over the head and body,
head wide, width less than to slightly more than length, large snout tubercle, two rows of tubercles from dorsal rows extend onto temporal and occipital areas, no occipital groove, chin barbel present,
dermal process behind jaw angle very large (base length 20-26% head length), rounded or scalloped in larger fish, first dorsal fin spines 3-4, no dark annular bands.
Morphology
There are 3-4 neuromasts in row 7 (shared with B. leobergius, usually one in other Benthophilus). The head is very wide , 103-111% head length (only B. leobergius has similar
measurements). First dorsal fin with 2-4 spines, usually 3, second dorsal fin with 1-2 spines, usually 1, followed by 6-10 soft rays, mostly 8-9. Anal fin with 1 spine followed by 6-9, usually 7, soft
rays. There are no evident gill rakers on the arch but rounded tubercles are present. Total vertebrae number 27-28, with 17-19 caudal vertebrae, usually 18. The head and body are covered with
minute granules with larger stellate scutes or tubercles bearing sharp spines on the body and on the dorsal and lateral head surfaces. Granules extend onto the medial half of the eyeball and onto the
pectoral fin base. Scutes are few and scattered on top of the head, strongly developed and close together on the side of the head. There are three longitudinal rows of tubercles on the body with
19-27, usually 22-23 (or 23-27, modally 25, authors differ) tubercles in the dorsal row, 8-16, usually 10-13, laterally, and 19-24 abdominally, usually 21-22. The ventro-lateral row is
usually absent. The belly is naked. The gut is moderately elongate with several convolutions.
Iranian specimens had the following meristics: first dorsal fin spines 3(3), second dorsal fin with 1 spine and 8 soft rays (4), anal fin with 1 spine and 7 soft rays (4), and pectoral fin with 16(1)
or 17(3) branched rays. The extent of tubercle rows is difficult to distinguish since their size gradually decreases posteriorly and their arrangement is irregular anteriorly. Dorsal row tubercles
25(4), lateral row tubercles 9(1), 13(2) or 14(1), and lower row tubercles 21(1), 22(2) or 23(1).
Sexual dimorphism
Mature males lose the granules, scutes and tubercles, their dorsal and anal fins enlarge and their cheeks become more muscular or inflated.
Colour
Overall colour is ash-grey with scattered melanophores on the upper flank and dorsal surface. Melanophores line the pectoral and caudal fin rays being best developed on the more dorsal rays.
Size
Reaches 13 cm.
Distribution
This species is reported from the Caspian Sea including the south from the Anzali Mordab and Gorgan Bay in Iran and between Kultuk and Astara in Azerbaijan (Ragimov, 1965).
Zoogeography
A Caspian Sea endemic.
Habitat
Found mostly in the sea over muddy bottoms but may approach river mouths. Commonly at 0.5-10.0 m, migrating to deeper areas at 20-25 m in winter. De Filipii (1865) reports this species from the Murdab
(= Anzali Mordab presumably) in relatively fresh water.
Age and growth
Young fish taken in early June are 2.4-3.5 cm and 0.35-1.00 g and by the end of November they are 7.0-8.0 cm and 10.15 g, the maximum values for the species. By February-March they are nearly mature
(Ragimov, 1985a). There are up to 4 age groups (Reshetnikov, 2002) although Pinchuk et al. in Miller (2004) suggest 18 months.
Food
Food includes molluscs (80%) crustaceans, fish, worms and chironomids (Zenkevitch, 1963). Iranian fish contained clams, crustaceans and a polychaete in gut contents.
Reproduction
Spawning takes place in April-May and rarely up to mid-June near the shore on silt or silt-sand bottoms with a mixture of shells. The eggs are laid in large, empty mollusc shells. Egg laying occurs in
two batches as females carry eggs of two sizes. Fecundity of the larger eggs is up to 3180 and the smaller 1403. Their diameters are 2.0-3.0 mm and 0.3-0.6 mm respectively. Young are caught in
early June (Ragimov, 1985a). Males probably die after spawning. An Iranian fish, 77.9 mm standard length, caught on 13 March had minute but developing eggs.
Parasites and predators
None reported from Iran.
Economic importance
None.
Conservation
Numbers in Iranian waters and threats to this species need to be assessed.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
Iranian material: CMNFI 1970-0543, CMNFI 1970-0544, mmm standard length, ();
CMNFI 4, 55.3-82.8 mm standard length, Gilan, Caspian Sea near Bandar Anzali ().
Benthophilus mahmudbejovi Endemic to the Caspian Sea and found on the western coasts of the Middle and South Caspian and the Volga delta region. No Iranian record.
Benthophilus pinchuki Common names
None.
[Pugolovka Pinchuka or Pinchuk's tadpole goby in Russian]
Systematics
Originally described as a subspecies of B. ctenolepidus, Pinchuk and Miller in Miller (2004) recognise it provisionally as a distinct species. This taxon was described from "raione Belogo
bugra y vostochnogo poberezh'ya Yuzhnogo Kaspiya" (= area of the White Hill (? locality not ascertained further by me) near the eastern coast of the southern Caspian Sea). A specimen listed as a
syntype is in the Natural History Museum, London (no number, noted in October 1982). The holotype of pinchuki is in the Caspian Biological Station, Institute of Zoology, Baku, Azerbaijan under
CBSIZA 52 (later in ZISP 53569, ex IZA), paratypes are under CBSIZA 304 (24, later 8 fish and ZISP 53660 9ex IZA 304) (6)). Additional material is in the Zoological Institute, St. Petersburg under ZISP
33143 (6) and 44346 (1) (Eschmeyer et al., 1996; on-line version downloaded 22 August 2007).
Key characters
Tubercles are vertically elongated, curved and crest-like, with a spiny fringe on their rear edge, dorsal row tubercles complete and 30-33, ventral row tubercles 23-27, granules are absent
postorbitally on the head and on the body, temporal and occipital region without large tubercles, head narrow, the interorbit has a shallow median groove, dermal filaments present or absent, chin barbel
usually present, dermal process behind jaw angle narrow (less than eye diameter) and barbel-like, first dorsal fin spines 3-4, and colour without dark bands.
Morphology
The first dorsal fin has 3-4 spines, usually 4, the second dorsal fin 1 spine and 9-11 soft rays, and the anal fin 1 spine and 8-11 soft rays (Ragimov, 1982; note the holotype has a much lower count
of anal fin rays than figure in table). Dorsal row tubercles 30-33, usually 31-32 (other species having 30 or less usually except B. ragimovi), ventral row tubercles 23-27, and dorso-lateral row
tubercles 10-23. Total vertebrae 30-32 and caudal vertebrae 21-23 (26-31 and and 17-21 in most other Benthophilus). The pelvic fin extends to the anus or beyond. Chin barbel less than half eye
diameter. One neuromast in row 7.
Sexual dimorphism
Males may have longer pectoral fins than females and grow larger than females.
Colour
Not described except for the absence of dark bands, blotches and spots. Probably an overall greyish.
Size
Reaches 8.9 cm in males.
Distribution
Found throughout the Caspian Sea.
Zoogeography
Endemic to the Caspian Sea.
Habitat
Known down to 282-294 m, usually 50-100 m but as shallow as 20 m, and a salinity of 12.4-13.2‰.
Age and growth
Life span is probably 18 months with maturity at 1 year.
Food
Unknown.
Reproduction
Spawning probably occurs in spring as fry have been caught in late April.
Parasites and predators
Unknown.
Economic importance
None.
Conservation
Numbers in Iranian waters and threats to this species need to be assessed. Boldyrev and Bogutskaya (2007) call it a rare deepwater species.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
No specimens examined, based on Pinchuk and Miller in Miller (2004).
Benthophilus ragimovi Endemic to the Caspian Sea from the western coast of the Middle and South Caspian, south to Astara. No Iranian record.
Benthophilus spinosus Endemic to the Caspian Sea from the eastern and western coast of the Middle Caspian , the southeastern North Caspian and southeastern South Caspian. No Iranian record.
Benthophilus svetovidovi A Caspian Sea endemic found on the eastern coast of the Middle Caspian, and
from the
south Caspian Sea by Naseka and Bogutskaya (2009). No Iranian record.
Genus Boleophthalmus This genus of gobies comprises several species in the Indian Ocean including one reported from Iran.
The genus is characterised by a very elongate, compressed body, a
rounded head, greatly thickened head and nape epidermis (as protection against
drying when out of water), scales cycloid, moderate to minute in size, eyes high on the head
and close together, lower eyelid or dermal cup well-developed, mouth slightly
oblique, teeth in a single row in each jaw, upper jaw teeth conical
with some canines, lower jaw teeth flattened, obliquely notched and
almost horizontal, tongue truncate and nearly all adnate to mouth
floor, gill opening small and oblique, and second dorsal and anal fins long (23 or more rays).
Boleophthalmus dussumieri
Common names
eshlambo, neeshlambo, gel khorak.
[kelb-el-daw, abou-shlembo, abu-shelamboo or triton in Arabic; gullo in Pakistan; Dussumier's mudskipper].
Systematics
Boleophthalmus Dussumieri was originally described from
Bombay, India and the holotype is in the Muséum national d'Histoire
naturelle, Paris under MNHN A.1468 (Bauchot et al., 1991).
Synonyms are Boleophthalmus dentatus Valenciennes in Cuvier
and Valenciennes, 1837, described from Bombay and reported from the
Shatt al Arab, Iraq by Iraqi authors, and Boleophthalmus chamiri
Holly, 1929 from "Chamir an der Straße von Clarence gegenüber der Insel Tawilah in der Meerenge von Hormus,
südöstliches Persien" (Holly, 1929a; Berg, 1949) (Khamir is at 26°57'N, 55°36'E in
Khowran Strait (= Clarence Strait) opposite Qeshm Island (= Tawilah or
Tavileh Island). Boleophthalmus boddarti (Pallas, 1770) as
reported by Relyea (1981) does not occur in the Persian Gulf region (Murdy, 1989).
Mirza et al. (1996) separate B. dentatus and B.
dussumieri on the basis of first dorsal fin height, body depth,
number of teeth on the jaws and the membrane between the two dorsal
fins. Murdy (1989) defines the genus Boleophthalmus in part by
the dorsal fins being separate, but does not give first dorsal fin
height, body depth and tooth counts for B. dussumieri or its synonyms.
A syntype of Boleophthalmus chamiri is in the Naturhistorisches Museum Wien (NMW 13804).
Key characters
The anal fin base and second dorsal fin base are 34% or more of
standard length and there are 2 canine teeth internal to the lower jaw
symphysis in contrast to Periophthalmus, the other mudskipper of southern Iran.
Morphology
First dorsal fin spines 4-5, usually 5, second dorsal fin with 1
spine and 24-28 soft rays, anal fin with 1 spine and 23-27 soft rays,
and pectoral fin branched rays 16-20. Scales are minute and embedded,
103-185 in longitudinal series; predorsal scales 48-56. Each scale is
rounded with radii around a posterior focus. Circuli are few. There
are 3 canine teeth on each side of the upper jaw (1-2 may be lost),
overlapping it when the mouth is closed and obviously protruding in
some preserved fish, and 39-76 upper jaw teeth and 43-71 notched lower
jaw teeth, the latter projecting almost horizontally from the jaw. The
number of teeth increases with age and B. chamiri was founded
on juveniles with fewer teeth (Berg, 1949). Gill rakers are partly
embedded in a membrane and not always easy to count; the range is
12-14 for Iranian specimens. Upper arm rakers are long and pointed,
less developed on the lower arm. When appressed, each raker touches
the adjacent one. Gut very elongate and coiled. Muscle blocks may be
evident in preserved fish.
Meristic values for Iranian specimens are:- first dorsal fin with 4(2) or
5(20) spines, second dorsal fin soft rays 24(3), 25(5), 26(9), 27(3),
or 28(2), anal fin soft rays 23(3), 24(4), 25(13), 26(1) or 27(1), and
pectoral fin rays 17(3), 18(8) or 19(11). Total vertebrae 26(10).
Sexual dimorphism
The male has a higher dorsal fin and larger canine teeth than the
female. Females were described as B. dussumieri while males
were described as B. dentatus (Berg, 1949).
Colour
Overall colour a greyish violet or light blue. The eye is light
yellow to brown around the rim. The opercle, pectoral fin base and
area just above the pectoral base have small green spots. There are 3
greenish-brown blotches on the nape, 3 more at the first dorsal fin
base and 2 others at the anterior second dorsal fin base. The first
dorsal fin is purple to light blue with small black or brown spots, or
with greenish spots at the base and wavy white lines and bluish spots
over the whole fin. The second dorsal fin has 2-6 horizontal rows of
oblong blue (or white in preservative) spots on the fin membranes on a
purplish background. Large white spots are found on the base of
interradial membranes 7, 10, 14, 17, and 22 on a specimen examined by
Murdy (1989) but this varies markedly between individuals seen by me.
The margin of this fin is black, edged with orange (white in
preservative). The caudal fin is black distally and bluish near the
base. The anal fin is transparent, the pectoral fin bluish and the
pelvic fin whitish.
In preservative, larger fish are darker than small ones. The body
is speckled darker above than below but this is still much lighter
than the dark dorsal fins. The first dorsal fin is very dark, almost
black, because the small pigment spots are so dense. The second dorsal
fin is lighter with alternate horizontal light and dark rows on the
membranes. Rays are somewhat lighter or equally dark. The second
dorsal fin margin is transparent but some fish may lack this feature
and be dark interradially (because of a dark row) and light at ray tips. The
caudal fin rays are dark and the membranes are dark next to the rays
but light in the centre. The anal fin is immaculate and the pectoral
and pelvic fins are very light with some pigment lining the rays. Some
pectoral fin pigment appears to be extensive vascularisation.
Peritoneum dark brown to black.
Young fish have several vague bars posteriorly on the upper flank
above the anal fin level. The anterior flank and side of the head is
covered with small spots.
Size
Attains 24 cm and 35.4 g.
Distribution
Found from Iraq to Pakistan and the west coast of India. Reported
from the Shatt al Arab, Iraq near Iranian Khuzestan by Menon (1956),
Khalaf (1961), and Al-Nasiri and Hoda (1976). In Iran it is recorded from the
Sarbaz River at Bahu Kalat in the Makran, and the Karun River near Mohammerah
(Berg, 1949) and in the Gulf, Hormuz and Makran basins.
Abdoli (2000) maps this species from the lower Karun and Arvand rivers in the Tigris River basin,
the lower Hilleh and Mand rivers in the Gulf basin, the Mehran, Kul and Minab
rivers in the Hormuz basin and lower reaches of Makran rivers from the Jagin to Bahu Kalat.
Zoogeography
This species has ready access to marine waters and can easily move between rivers via the sea.
Habitat
This species is active on mud flats in streams under and above
tidal influence but most information on behaviour and biology is based
on studies of fish on marine tidal flats (as recounted below). In
southern Iranian rivers, this species can be seen swimming with its
head out of the water and pectoral fins splayed, presumably on the
look-out for danger. Populations are very numerous on mudflats on
lower reaches of rivers in Khuzestan. It hides in mud burrows when
approached and a plume of dark muddy water can be seen emerging from
the burrow, or it may bounce over the water surface. They are
difficult to catch with a seine when approached from the shore because
of this behaviour, scurrying rapidly from the land and shallows into
deeper water, muddying the water and obscuring vision, and thus
disappearing from view. However they can be caught in shallower rivers
by using a seine net from mid-river to shore, after waiting for them
to re-emerge and enter the shallows. Their reaction is still flight to
water but the movement from water to land confuses them and they can
be scooped up in their panic. In Khuzestan, local people whip them out
of the water with palm fronds.
Both sexes of this species construct U-shaped, vertical burrows in
sand to a depth of about 1 m in marine situations. Clayton and Vaughan (1986)
illustrate burrow shapes, outline construction methods and experimented on
territoriality by removal of fish. A polygonal, mostly pentagonal but
significant numbers of hexagonal and quadrangular, dam of mud is built up around
the burrow hole using the mouth, again by both sexes, and is used to reduce
aggression between neighbours (Clayton
and Vaughan, 1982; Clayton and Wright, 1989). This dam serves to keep water in the burrow at low tide
although not all populations maintain this dam as completely. The burrows are
used as shelter from low temperatures during winter and high temperatures in
summer, at high tides, and when danger threatens. Air temperatures in Kuwait
reach 51.1°C in summer and fall to -4°C in winter, conditions mirrored in Iraq.
Polygonal dams were not seen in rivers, perhaps because of river flow and tidal
fluctuations. Fish are active in the 13-40°C range and daily variations may be
as much as 20C°. Fish tend to be inactive in winter in the northern Gulf, with
feeding the main activity, and are easy prey for migratory birds feeding on the mudflats.
At low tide the fish emerges and raises its head, scanning the
surrounding surface and recognising humans as threats at 5-7 m.
Movement is with the aid of the muscular pectoral fins and the gill
covers are puffed out, presumably to obtain water or to maintain a
moist atmosphere for the gills. It does not usually travel further
than the mud walls of its territory in contrast to Periophthalmus
waltoni and is exposed to air for much shorter periods being
submerged in water or watery mud most of the time. The goby basks when
recently emerged from its burrow in order to raise body temperature to
14°C before moving out onto the mud. The minimum temperature for emergence
is 10°C. Some larger fish will make rapid and short sorties from the burrow to
test air temperatures. Evaporative loss of body fluids is used to cool
the fish in summer and the body is kept wet by immersion in pools and
burrows. Body temperatures do not exceed 33°C even when air temperatures were over 40°C.
There can be up to 52 fish in 100 sq m in Kuwait (Tytler and Vaughan
(1983) and Clayton and Vaughan (1988) on B. boddarti in Kuwait
marine waters, identified as this species by Murdy (1989)). At the Khowr-e Mussa
in Iran, each fish defends a dammed territory of about 1 sq m, this area
containing the epiphytobenthic food it requires (Höpner and Kazem Maraschi, 1999).
The skin has large numbers of mucous cells which prevent desiccation on the mud
flat and lubricate the fish during burrowing (Salih and Al-Jaffery
(1980) on B. dentatus from Al Faw (= Fao), assumed to be this species). Gas
exchange in air and water takes place through the gill, inner
operculum, nasal, body and outer operculum skin in that order, as
adaptations to low oxygen conditions in estuarine areas and to life
out of water (Al-Kodhomiy and Hughes (1988) on B. boddarti but
presumed to be this species). In addition to the territorial
individuals, there are also smaller, non-territorial fish which roam
more freely and are called "errant" (Clayton and Vaughan,
1988). Clayton and Vaughan (1988) describe in detail a series of body
postures and movements associated with being stationary, moving,
feeding, maintaining moisture levels, construction, interacting with
other individuals, avoiding predators, and courtship. Males raise
their dorsal fins when they meet, gape widely and fight mouth to mouth.
Age and growth
On the Karachi coast of Pakistan, juveniles and young of the year
are found from September to December, growing from 45 mm standard
length to 60 mm over three months (Hoda and Akhtar, 1985). In Gujarat,
India three age groups are reported (Soni and George, 1986).
Food
Diet includes diatoms (Pankow and Huq (1979) recorded 114 species at Al Faw (= Fao)),
blue-green algae, filamentous algae, insects, crustaceans, nematodes and teleost
fish eggs in marine situations, the polygonal enclosure acting as a
"farm" promoting growth of diatoms (Sarker et al., 1980;
Clayton and Vaughan, 1988). It grazes on surface films of yellow-green algae at
Fao (Pankow and Huq, 1979) and brown algae in Kuwait. The grazing action,
with side-to-side head movements, is distinctive for this species
among other mudskippers in the Gulf (Clayton and Vaughan, 1988). The
fish leave their burrows on feeding excursions. Young are found in
non-territorial groups in shallow pools, where in winter they take
some time to warm up before leaving to feed. The pools offer a
protective environment for the young.
These mudskippers are fed on by gulls, terns, bitterns, egrets and
herons as well as a wide variety of other fishes and sea snakes (Clayton and Vaughan, 1988).
Reproduction
Spawning in the marine environment of the Khawr az Zubayr, Iraq takes place in June,
the release of larvae being timed to take advantage of phytoplankton and
zooplankton blooms (Hussain and Ahmed, 1999). On the Karachi coast of Pakistan, 50% maturity (= first maturity)
is given as 69 mm standard length in males and 72 mm in females (Hoda
and Akhtar, 1985) while maturation starts at 60-65 mm standard length
in males and 70-75 mm in females according to Hoda (1987). Spawning
there occurs twice each year during April-May and July-September (Hoda,
1986) or April-May and July-August (Hoda and Akhtar, 1985). Fecundity
range is 986-4912 eggs for fish 74-110 mm long and maximum egg
diameter is 1.05 mm (Hoda, 1986). Hoda and Akhtar (1985) give a range
of 970 eggs at 75 mm standard length to 4113 eggs at 110 mm, mean 2371 eggs.
Clayton and Vaughan (1988) give the start of the breeding season in
Kuwait as March but the majority of males are only displaying by
mid-April. Courtship continues until August and most territories have
breached walls where fish have crossed over into other territories to
find mates. Larvae appear as early as mid-July although most appear
later than this. Territories are repaired in October and November.
Courtship involves competition between males for neighbouring
females. Each male has on average 5 neighbours but only half will be
females. Males identify themselves and try to attract females by
elevating their fins and leaping from a lateral prone position with
about two-thirds of the fish clearing the ground. Other signaling
devices are pectoral fin waves, lateral tail-beats, and quivers. The
female advances and the male leads her into his burrow where spawning
presumably occurs. While errant females may be mated with territorial
males, errant males having no territory do not show the leaping behaviour.
Parasites and predators
Migrating birds are heavy predators of this fish on mud flats (Tytler
and Vaughan, 1983). Indian pond herons are known to eat this fish in
Iran. E. Kahrom in the Crocodile Specialist Group Newsletter reports that
muggers (marsh crocodile, Crocodylus palustris) eats a Periophthalmus
in the Sarbaz River of Iranian Baluchestan (www.flmnh.ufl.edu/natsci/herpetology/newsletter/news173b.htm,
downloaded 15 February 2002). However this may well be Boleophthalmus dussumieri.
Economic importance
None, although children play with them in the Karun River
(www.abadan.net/abadanidictionary.html, downloaded 4 December 2003).
Conservation
There has been no assessment of the conservation status of this species but
it is common on mudflats in the lower reaches of rivers Iran.
Further work
The biology of this species in Iranian fresh waters has not been investigated.
Sources
Iranian material: CMNFI 1979-0141, 1, 86.2 mm standard length, Hormozgan, Rud-e Kul (27º17'30"N, 56º03'30"E);
CMNFI 1979-0142, 5, 74.6-137.5 mm standard length, Hormozgan, Baghu River (27º17'N, 56º28'E);
CMNFI 1979-0145, 10, 42.6-136.6 mm standard length, Hormozgan, Geru River (26º55'N, 57º01'30"E);
CMNFI 1979-0322, 5, 57.1-111.8 mm standard length, Baluchestan, Bahu Kalat River (ca. 25º45'N, ca. 61º26'E);
OSU 8121, 1, 61.4 mm standard length, Baluchestan, Bahu Kalat (no other locality data).
Comparative material:- CMNFI 1985-0182, 3, 44.7-68.1 mm standard length, Iraq, Shatt al Arab (no other locality data);
BM(NH) 1976.12.8:4-7, 4, 103.9-128.2 mm standard length, Iraq, Basrah (30º30'N, 47º47'E);
BM(NH) 1954.11.10:3-7, 6, 79.9-147.5 mm standard length, Iraq, Fao (29º58'N, 48º29'E).
Caspiosoma caspium
Caspian Sea basin, described and recorded from the northern and
middle Caspian Sea, but no Iranian record. Ragimov (1998c), Miller (2004) and
Reshetnikov (2002) give recent descriptions.
This genus has a single species in the
Caspian Sea. The characters of the genus are covered in more detail by Miller in
Miller (2004) and are the characters of the species below. It is recognised as
distinct from the related Neogobius by head sensory papillae patterns
(especially an additional transverse row before row b) and modally 7
first dorsal fin rays rather than 6. The number of the transverse suborbital neuromast rows of the cephalic lateral
line system is an important diagnostic feature for the monotypic genus Chasar
according to Miller (2004). Pinchuk and Ragimov (1985) count these rows consecutively from anterior to posterior as
1 – 8 while Miller (2004) postulates an additional transverse row anterior to
the longitudinal suborbital row b labelling the posteriormost row as row
7 (Pinchuk et al. in Miller, 2004). A Farsi common name for Neogobius
and related gobies is گاو ماهي (gav mahi).
Chasar bathybius
Eichwald, 1831
Ragimov, 1978
Kessler, 1877

Ragimov, 1978
Kessler, 1877

Kessler, 1877
Kessler, 1877
Berg, 1927
Berg, 1949

Kessler, 1877
Kessler, 1877
(Pallas, 1788)

Ragimov, 1976
Ragimov, 1982
Boldyrev and Bogutskaya, 2004
Kessler, 1877
Pinchuk and Ragimov, 1979
Valenciennes, 1837
Valenciennes, 1837

(Kessler, 1877)

(Kessler, 1877)


Common names
[bychok-glubokovodny or deepwater goby]
Systematics
Described from Svinoi Island south of Baku, Azerbaijan. No types known. Neilsen and Stepien (2009) include this species in Ponticola in their molecular analysis of endemic Ponto-Caspian gobies but, as they did not include any material of this species nor any discussion of the nominal genus, it is left in Chasar here.
Key characters
Nape with cycloid scales anteriorly, not quite reaching orbit level, head depth at eyes about equal or slightly less than width between upper origins of opercles, interorbital distance equal to or slightly less than orbit, angle of jaw below pupil of eye, snout longer than eye, up to 1.8 times orbit diameter, upper lip not swollen at angle, pelvic fin not reaching anal fin, except in young, anterior pelvic fin membrane without well-developed lateral lobes, caudal peduncle depth 0.3-0.5 length, first dorsal fin with 7 rays and little higher than second dorsal fin (which is lower posteriorly), overall colour semi-transparent, and lateral series scales mostly 55-60.
Morphology
First dorsal fin with 6-8 spines, usually 7, second dorsal fin with 1 spine and 14-16 soft rays, and anal fin with 1 spine and 11-14 soft rays, lateral series scales 55-65. Scales are overall ctenoid but cycloid on the nape, breast, opercle, abdomen and and lobe of the pectoral fin. The pectoral fin is long reaching back well beyond the level of the second dorsal fin origin. Lateral line system usually with eight or nine transversal suborbital neuromast rows, two rows ventral to the longitudinal suborbital row b. Four neuromast rows on the opercle. Lateral line system with eight suborbital neuromast rows arranged transversally, five before, three above and two below the longitudinal row b. Four rows of free neuromasts on the opercle instead of the characteristic three rows of all other neogobiines.
Meristics for Iranian specimens: first dorsal fin with 6(1), 7(14) or 8(1) spines, second dorsal fin with 14(4), 15(9) or 16(3) soft rays, anal fin with 11(3), 12(9) or 13(4) soft rays, pectoral fin with 18(10) or 19(6) rays, scales in lateral series 55(1), 56(3), 57(6), 59(1) or 60(1), and transverse rows of scales 16(8) or 17(4).
Sexual dimorphism
Males grow larger than females.
Colour
The body is semi-transparent with reddish or yellowish-brown colour. The myotomes are visible but scales are difficult to see. The back is darker than the flank and belly. Preserved fish have 4 faint saddles and faint spots along the lateral midline, the first dorsal fin has two brownish bands, and young have a stripe-like spot posteriorly on the first dorsal fin.
Size
Attains 29.0 cm.
Distribution
Caspian Sea basin, recorded between Kultuk and Astara in Azerbaijan (Ragimov, 1965). Kiabi et al. (1999) report this species from the southeast Caspian Sea, southwest Caspian Sea and south-central Caspian Sea in Iran. Miller in Miller (2004) maps this species from the Safid River westwards and from Gorgan Bay but there are no text records to go with the map. Recently recorded from Iran (Ahnelt, et al., 2007) and the Anzali Shore (K. Abbasi - see photograph above). South coast of the Caspian Sea especially in Gorgan Bay (Jolodar and Abdoli, 2004).
Zoogeography
Endemic to the Caspian Sea but related to the speciose genera Neogobius and Ponticola.
Habitat
A marine species, not entering fresh waters, found down to 75 m and perhaps deeper, especially in colder conditions. It is found on sandy and shelly bottoms and in smaller numbers on firm silt.
Age and growth
Life span is estimated to be 4 years, with maturity in the second year and spawning every year. Abdoli et al. (2006) examined fish from the Gomishan and Miankaleh wetlands and found a length (L)-weight (W) relationship of W = 0.0777L2.44 for sexes combined.
Food
Fish are an important food for larger specimens, smaller fish take crustaceans. Fish food includes Knipowitschia iljini and crustaceans include the decapod Palaemon, corophiids, mysids and some gammarids. A small number of worms (Nereis) are also eaten.
Reproduction
Eggs of only one size have been found in this species, suggesting a single spawning each year. Up to 2979 eggs have been recorded with a maximum diameter of 2.6 mm. Spawning in the southern part of the Middle Caspian Sea takes place from the second half of June until July. Males approach the coast in the first half of March or April while females remain at 20-30 m. Females enter shallow water in late April and in greater numbers in May and June. After spawning, fish retreat to areas deeper than 10-15 m.
Parasites and predators
Eaten by sturgeons and the Caspian seal. Sattari et al. (2004; 2005) surveyed this species in the inshore area of the Caspian Sea, recording Anisakis sp. and Dichelyne minutus.
Economic importance
None in Iran but caught in Lenkoran, Azerbaijan with fixed nets and in northern Azerbaijan with cast nets. The fish are oily but pleasant tasting (Pinchuk et al. in Miller, 2004).
Conservation
Kiabi et al. (1999) consider this species to be data deficient in the south Caspian Sea basin according to IUCN criteria. Criteria include medium numbers, medium range (25-75% of water bodies), absent in other water bodies in Iran, and absent outside the Caspian Sea basin.
Further work
The distribution of this species in Iranian waters needs clarifictaion.
Sources
Based on Pinchuk et al. in Miller (2004) and Ahnelt et al. (2007).
Iranian material: CMNFI 1971-0352, 8, 141.7-195.8 mm standard length, Gilan, Caspian Sea + Shalman River (37°28’N, 49°27'E, 37°08’N, 50°15’E); CMNFI 2006-0022, 3, 142.9-195.4 mm standard length, Gilan, southwest Caspian Sea off Astara (38°00’N, 49°30’E to 38°20’N, 50°00'E); CMNFI 2006-0023, 1, 99. 4 mm standard length, Gilan, Chaboksar (36°58’N, 50°34’E); CMNFI 2006-0024, 2, 160.4-183.1 mm standard length, Gilan, Talesh (37°48’N, 48°55’E); CMNFI 2006-0025, 2, 174.7-187.9 mm standard length, Gilan, Talesh (37°48’N, 48°55’E); CMNFI 2006-0026, 2, 76.7-81.3 mm standard length, Gilan, southwest Caspian Sea off Bandar Kiashahr (37°25’N, 49°57’E).
The gobies of this genus are found in fresh, brackish and marine waters in the Indo-West Pacific area. Most species are freshwater residents and there are about 24 species with one in Iran. They are characterised by an elongate body, anteriorly rounded and posteriorly compressed, head depressed, snout elongate, scales on the body ctenoid and on the head cycloid, head scaled behind eyes, cheeks and operculum scaleless, anterior nostril a short tube, lower jaw prominent, tongue bilobed, teeth in several rows in each jaw, gill openings wide, isthmus narrow, no flaps on the inner edge of the shoulder girdle, and pectoral fin without free rays and base scaled.
Glossogobius giuris
(Hamilton, 1822)




Common names
gel-ye mahi cheshm navari (= band- or bar-eyed goby) or gavmahi-ye cheshmnavari, rikoo (sand dweller or benthic fish, local name courtesy of Asghar Mobaraki).
[guloo in Pakistan; bar-eyed goby, tank goby, white goby, flathead goby, crocodile goby].
Systematics
Gobius giuris was originally described from the Ganges River, India.
Key characters
As the sole representative of the genus in Iran, the characters of the genus serve to identify it.
Morphology
The first dorsal fin has 6 spines, the second dorsal fin has 1 spine followed by 7-9 soft rays, the anal fin has 1 spine and 7-9 soft rays and the pectoral fin has 16-21 branched rays. Lateral series scales 28-36. Scales on the head are mostly cycloid while body scales are ctenoid. The exposed portion of flank scales are diamond-shaped. Scales are rectangular in overall shape with a rounded anterior margin which may be slightly indented above and below the mid-point, parallel dorsal and ventral margins, and a posterior margin with two straight edges meeting at a rounded central tip. The anterior dorsal and ventral corners are sharp. There are fine ctenii on the posterior margin. Numerous radii radiate anteriorly from the rounded central tip of the posterior margin. Circuli are fine. Esmaeili et al. (2009) give details of scale morphology using scanning electron microscopy. Gill rakers are short and reach the one below when appressed. The cheek has 3-5 longitudinal mucus canals. The gut is short and s-shaped.
The chromosome number is 2n=46, with 46 acrocentric chromosomes and 46 chromosomal arms (Subrahmanyam, 1969; Ráb, 1985; Vasil'ev and Grigoryan, 1993; Klinkhardt et al., 1995).
Meristics on Iranian specimens: first dorsal fin with 6(3) spines, second dorsal fin soft rays 9(3), anal fin soft rays 8(3), and pectoral fin rays 19(1) or 20(2). Scales in lateral series 32(1) or 36(1), total gill rakers 12(1) or 14(1) and total vertebrae 27(1).
Sexual dimorphism
Males are larger than females. Male dorsal fins are longer and their overall colour is brighter (Pethiyagoda, 1991).
Colour
Olive-green to blackish-green or yellowish to sandy brown on the back with flanks similar but lighter, often a greenish-yellow. The belly is cream coloured. The head and body are darkly blotched and spotted. The flank may have 5 regularly spaced blotches. The sides of the head have irregular dark to violet spots. The fins have thin bars appearing as rows of spots on each fin ray, in some concentrated at the base of the second spine of the first dorsal fin. Fins are a yellowish-green. The first dorsal fin may have a dark spot posteriorly and the anal fin a black margin. The pectoral fin has a dark spot above the base. Small preserved fish are immaculate. The peritoneum is silvery with a few small and scattered melanophores.
Size
Attains about 50 cm.
Distribution
Found in the Indo-West Pacific entering rivers. Abdoli (2000) maps this species as questionably present in the lower Karun and Arvand rivers of the Tigris River basin, questionably in the Gulf and Hormuz basins; and in lower reaches of Makran rivers from the Jagin to the Bahu Kalat or Sarbaz. It is recorded from the Hormuz and Makran basins by specimens.
Zoogeography
The marine abilities of some members of this family would aid in dispersal between river systems connected to the sea.
Habitat
Sand and mud substrates are preferred over rock in Sri Lanka (Pethiyagoda, 1991). The fish lies embedded in the sand with only the eyes protruding and rarely swim freely. Young fish may form schools or may rest close to shore in shallow water where they can be caught by hand. Tekrival and Rao (1999) report its aquarium preferences and habits as 22-25°C, pH 6.5-7.2, predator, almost dark lighting, bottom dwelling with stones preferred and a pair or more can be kept. Asghar Mobaraki (pers. comm., 8 January 2012) recorded a fish kill of this species on 26 December 2011 at the Zirdan Dam construction site on the Kaju River.
Age and Growth
Hossain et al. (2009) found a value close to isometric for this fish in the Ganges River of northwestern Bangladesh (b = 3.068 for total length and combined sexes, 2.954 for males and 3.293 for females)
Food
Diet includes small fishes seized from its concealment in the sand by a sudden lunge; this species being a visual ambush predator. It may browse on the substrate and take living food. An Iranian specimen had crustacean remains.
Reproduction
Eggs are green and when laid become strongly attached to any submerged structure by a tube 3-8 mm long. The eggs are 0.5 mm in diameter. Freshwater populations may migrate to the sea to breed as there is a marine larval stage although some are reported to be land-locked in South Asia and hence freshwater breeders (Pethiyagoda, 1991). The spawning period is June to September, peaking in July to August (Saksena, 1980).
Parasites and predators
None reported from Iran.
Economic importance
This goby is a food fish in South Asia and can be caught on light angling gear. It has been reported as being ciguatoxic (intermittently poisonous through feeding on toxic food) (Bagnis et al., 1970).
Conservation
The distribution and abundance of this species in Iranian waters has not been examined and no conservation assessment can be made.
Further work
This species seems to have a fairly restricted distribution in Iran and this should be investigated further.
Sources
Iranian material: CMNFI 1979-0322, 2, 20.7-53.6 mm standard length, Baluchestan, Sarbaz River (ca. 25º45'N, ca. 61º26'E);
OSU 8124, 1, 162.0 mm standard length, Baluchestan, Bahu Kalat, Sarbaz River (25º43'N, 61º25'E).
Genus Hyrcanogobius Hyrcanogobius bergi This
species was described from the north Caspian Sea near the mouths of the
rivers Volga, Ural and Emba. Ragimov (1977) reports this species from off Gasan
Kuli at 37°30'N in Turkmenistan near Iranian waters. Shakirova and Sukhanova (1994) record this
species from the Atrek lakes of Turkmenistan, the Atrek River flowing into Iran
for part of its course.
Gobius longecaudatus var. c Kessler, 1877 described from Baku Bay
is a synonym.
Iljin, 1928
Iljin, 1928
Genus Knipowitschia
Iljin, 1927
This genus of sand gobies has about 16 species in the Caspian, Black, Aegean and Adriatic seas and adjacent fresh waters with 2 recorded from Iran. Later spelt Knipovitschia, incorrectly, in the English summary of Iljin's 1928 description. The original description is apparently in Iljin (1927d) (as is that of Caspiosoma) but was repeated in Iljin (1928). Hyrcanogobius Iljin, 1928 and Bubyr Iljin, 1930 are synonyms (Economidis and Miller, 1990) although Miller in Miller (2004) separates Hyrcanogobius.
The body is fusiform without a strongly depressed head, the dorsal profile anteriorly being straight. The mouth angle does not reach past the anterior part of the eye. The mouth is oblique. The eyes are moderate in size and may be lateral or directed upwards. The body usually bears imbricate ctenoid scales but the head is naked as can be the anterior part of the back as far as the second dorsal fin and the middle of the abdomen to the anal fin. There may be axillary scales and scales on the belly midline. Some members lack scales except for the axillary region and the caudal peduncle. Lateral series scales less than 40. There are no barbels and the anterior nostrils are not an extended tube-like structure. The posterior nostrils are pore-like. The nape lacks a bony crown and only the posterior part of the cranium is covered with dorsal muscles. The preoperculum lacks teeth or projections. The isthmus is broad and the branchiostegal membrane is attached to its entire lateral margin. Jaw teeth are conical, caniniform and in several rows forming a band. The outer row is slightly enlarged but there are no strongly enlarged teeth. The vomer lacks teeth. The tongue is truncated or slightly rounded. The gill rakers are small and broadly spaced apart. The pectoral fins lack fine, silk-like rays. The anterior edge of the pectoral girdle lacks dermal flaps. The pelvic fin disc reaches to, or near to, the anus. The pelvic fin anterior membrane is well-developed but without obvious lateral lobes. The caudal peduncle is slender and longer than the second dorsal fin base. The upper angle of the caudal fin is nearly rectangular and the caudal fin is rounded and shorter than the head. The head canals are variably developed or absent, and may be damaged by abrasion in poorly preserved material. The first dorsal fin modally has 6 or 7 rays and vertebral modes are 30-33. The swimbladder is present or reduced. There is no perianal organ. Colour is light. Postlarvae are planktonic.
Miller in Miller (2004) also defines this genus by characters of the head lateral line system, namely having the infraorbital longitudinal row a present with typically at least 2 transverse rows from this row; transverse infraorbital rows between levels of rows b and d along cheek, with rearmost descending behind posterior end of row d; transverse row behind jaw angle not descending below level of row d; row b short, to below rear eye; anterior row tra distant from row b; anterior dorsal row g distant from row n; snout rows r and s linear; uniserial row of papillae along upper edge of each orbit when canals are absent but never with transverse rows; anterior oculoscapular canal (if present) ending anteriorly in interorbit, dividing from pore κ to end at double pores β; posterior oculoscapular present or absent; and preopercular canal when present always lacks middl;e pore δ.
Vasil'eva and Kuga (2001) propose that the separation of Knipowitschia from Pomatoschistus is doubtful based on similar cranial osteology and the variation in the degree of development of head canals seen in Knipowitschia. However Miller in Miller (2004) disagrees and presents arguments for a "sand-goby" group of 5 distinct genera, of which only Knipowitschia and Hyrcanogbius are known from Iran (Pomatoschistus, Gobiusculus and Economidichthys being the others).
Miller (1990) states that the majority of the members of this genus evolved after the draining of the Western Paratethys basin into the Adriatic Sea basin in the late Miocene about 6.0-5.5MYA.
.Members of this genus are called gav mahi and سگ ماهي (sag mahi) in Farsi and this not repeated under each Species Account.
Knipowitschia caucasica
(Berg, 1916)


Common names
gel-ye mahi qafqazi (= Caucasian goby) or gavmahi-ye qafqazi.
[gafgaz xulu in Azerbaijan; bubyr kavkazskii in Russian; Caucasus bald goby; Caucasian dwarf goby].
Systematics
Pomatoschistus caucasicus was originally described from Batumi/Temirgoe, a station south of the Sulak mouth on the Black Sea.
The name of this species first appeared in Kavraiskii in Radde (1899) as a museum name without a description ( a nomen nudum) and was subsequently made available by L. S. Berg in 1916 with the type locality being a swamp near Batum and Lake Inkit near Pitzunda, Georgia (see Eschmeyer (1990) and Eschmeyer et al. (1996) for details).
Gobius lenkoranicus Kessler, 1877 described from "pribrezhnom' bolote, po blizosti Lenkorana" (= a coastal marsh near Lenkoran, Azerbaijan) is a synonym, tentatively so according to Economidis and Miller (1990). This name was suppressed by the International Commission on Zoological Nomenclature (see Kottelat, 1997; Miller et al. in Miller, 2004)). This earlier name for what could be caucasica was suppressed as a nomen dubium because of an inadequate original description in its details and the small and poorly-preserved holotype which could not be readily compared with caucasica. The name could also have been suppressed as a nomen oblitum as it had not been used for 50 years (Economidis and Miller, 1990).
Formerly placed in the genus Pomatoschistus Gill, 1863 and in the genus Bubyr Iljin, 1930.
Three syntypes from Lake Temirgorje, Georgia (formerly in the Tiflis Museum) are in the Natural History Museum, London under BM(NH) 1896.3.28:26-28 (22.1-25.2 mm standard length) (Eschmeyer et al., 1996; personal observations). Economidis and Miller (1990) list these three specimens (as evidenced by the catalogue numbers 26-28) as two males (22.8-24.0 mm standard length) and two females but give the length of only one female (24.5 mm). Miller et al. in Miller (2004) consider the material used by L. S. Berg (see above) to be the type series (ZISP 15343 from Lake Inkit and ZISP 15321 a swamp near Batum).
Key characters
The anterior oculoscapular lateral line head canals unite in the posterior interorbit, with a single median pore κ, and canals extend anteriorly to pores λ. The preopercular canal is present (Miller in Miller, 2004).
Morphology
First dorsal fin with 5-8, usually 6, spines (rarely 4 as an anomaly, e.g. in Berg (1948-1949, Fig. 784 (Iljin, 1956)), second dorsal fin with 1 spine and 6-10 branched rays, usually 7-8, anal fin with 1 spine and 5-10 branched rays, usually 7-8, and pectoral fin with 13-18 rays. Lateral scales 29-38. Vertebrae 30-33. The back in front of the first dorsal fin and the belly in front of the anal fin are usually scaleless. The caudal peduncle is completely scaled and there is a complete row of scales anteriorly along the lateral midline, although other flank scales may be missing. The anterior oculoscapular head lateral line canals join in the posterior interorbit region, with one pore κ, and paired canals in the interorbit each to a pore λ and single pore α. The posterior oculoscapular canal is typically present, the preopercular canal is present and there are no additional pores on the horizontal anterior oculoscapular canal behind the eyes (Miller et al. in Miller, 2004). The subopercular canal is variably present which has led to different generic assignments for this species. The larva has been described by Daoulas et al. (1993).
Meristic values for Iranian specimens are:- first dorsal fin spines 6(4), second dorsal fin rays after a spine 6(1) or 8(3), anal fin rays after a spine 8(2) or 9(2), lateral scales 33(1), 34(1), 36(1), 39(1), and total gill rakers 9(3) or 10(1). Rakers are very short, not reaching half way to the adjacent one when appressed.
Sexual dimorphism
See under colour. Males are larger than females. There are various morphometric differences (Miller et al. in Miller, 2004).
Colour
Overall colour is grey to fawn with olive-green tints. Fish in hypersaline habitats are darker. Males are generally darker than females and have distinct dark bars on the flanks below the two dorsal fins and on the caudal peduncle while females have irregular spots or blotches along the flank mid-line. Nuptial males have 4 prominent bars. However some males may lack bars and have spots and some females lack spots (Ahnelt et al., 1995). The head in males is densely pigmented, including ventrally. The caudal peduncle is completely pigmented, usually including the ventral surface. The flank bar under the first dorsal fin in males is long and usually runs from near the dorsal mid-line to the belly. The male first dorsal fin has three oblique dark bands and a dark spot medially on rays 5 and 6 extending posteriorly on the fin membrane. The distal edge of this dorsal fin is dark. The second dorsal fin has three bands also and a dark edge. Males have dark pigment on the pelvic fins, head and breast, and a silver hue on the anal fin during the breeding season. The male pectoral fin has a short and oblique dark band on the upper rays. The anal fin in males is normally dark with a pale margin while in females it is pale overall. The caudal fin has thin dark bands.
Females are a pale fawn overall. The female has a densely spotted first dorsal fin while the male has a blue blotch at the fin rear when in breeding condition. The first dorsal fin in females has a distinctive middle band but the others are faint. The second dorsal fin in females is only faintly banded. Breeding females have 2 intensely black areas on the first dorsal fin, a black spot on the lower jaw, 2 rows of melanophores running obliquely from the eyes to the upper jaw, and a yellow abdominal region (Economou et al., 1994). The back has fine melanophores extending to the rear of the second dorsal fin. There are light saddles at the origins of the first and second dorsal fins, the end of the second dorsal fin and on the caudal peduncle. The mid-flank has several blotches, some slightly deeper than wide. The rest of the body below these blotches is pale except for some short vertical groups of melanophores below the lateral midline and melanophores along the base of the anal fin. The breast is pale as opposed to dark in males.
Large and minute pigment blotches and spots variably developed are scattered over the whole head and body in both sexes of an Iranian sample. The peritoneum has large pigment spots ventrally or is almost immaculate.
Size
Reaches 48.6 mm, perhaps 69 mm, total length. Fish in Gorgan Bay reached 46.0 mm (Afraei and Hassannia, 1999).
Distribution
Found in the Aegean, Black and Caspian Sea basins. Introduced to the Aral Sea. Caspian localities include Imeni Kirova or Kyzylagach Bay near the northwestern Iranian border, the lower Safid River and lower Babol River, near Anzali and Ashuradeh (Derzhavin, 1934). It is reported from Gorgan Bay, the delta of the Karasu, the Atrak River, the Aras Dam the southeast Caspian Sea, southwest Caspian Sea and south-central Caspian Sea (Kiabi et al., 1999; Jolodar and Abdoli, 2004; Miller et al. in Miller, 2004; Abdoli and Naderi, 2009; K. Abbasi - see photograph above).
Zoogeography
This species is part of marine fauna in the eastern Mediterranean and the Black-Caspian-Aral seas basins and its relationships are outlined under the genus description above.
Habitat
This species lives and reproduces in salinities as high as 59.5‰ and up to 83‰ (Zenkevitch, 1963; Miller et al. in Miller, 2004) but also enters fresh water. Preferred conditions in the Aegean area are 15-20‰ (Ahnelt et al., 1995; Miller et al. in Miller, 2004). Juveniles prefer sand, mud or gravel bottoms near shore in Greece (Daoulas et al., 1993) but weedy shallows are also favoured (Economidis and Miller, 1990). They may overwinter in deeper water at 1-2 m in the Volga delta (Economidis and Miller, 1990) and migrate into shallow water in April-May prior to spawning (Miller et al. in Miller, 2004). The Atrek River population moves downstream (Miller et al. in Miller, 2004). Afraei et al. (2001) found this species to be most frequent in Gorgan Bay in January and least frequent in March.
Age and growth
Life span may be only 1 year and as a consequence numbers can fluctuate widely in any given year depending on environmental conditions. In the Evros Delta of the north Aegean Sea, fish grow rapidly during the summer after hatching, breed from the end of April to the end of July in the following summer and grow rapidly again from July to September. Older males die the following year after February but some females will survive to spawn a second time at the end of April and the beginning of May and then die. Fish may be mature at 14.5 mm total length (Kevrekedis et al., 1990). In the spawning season, females comprise as much as 85.5% of the population (Miller et al. in Miller, 2004).
A study of this species in Gorgan Bay, Golestan (Iranian Fisheries Research Organization Newsletter, 23:2, 2000) showed that the maximum size obtained was 46 mm with average lengths of 33 and 37 mm for males and females respectively. The sex ratio was 1:1.1 for males to females and growth is positive allometric (see also Afraei and Hassannia (1999) and Afraei et al. (2001)). In the Volga delta spawning males were 33.3 mm and females 30.2 mm on average (Miller et al. in Miller, 2004). Abdoli et al. (2006) examined fish from the Gomishan and Miankaleh wetlands and found a length (L)-weight (W) relationship of W = 0.135L2.72 for males, 0.0377L1.97 for females and 0.0182L2.51 for sexes combined.
Food
Diet was dominated by polychaete worms and chironomids (88%) in one study but crustaceans such as amphipods, copepods, ostracods, cladocerans, insects such as choronomids, bivalve mollusc larvae, and fish are also taken and items may be both benthic and planktonic.
Reproduction
Females may be carrying eggs at 20-24 mm and 8-10 months of age. Fecundity reaches 1389 eggs but can be as low as 60 eggs. Fecundity in the Caspian Sea ranges from 209 to 786 eggs with a mean of 423, from 209 to 382, mean 285 and from 527 to 863, mean 715.6 for various reports (Kevrekedis et al., 1990). Batches consist of 80-100 eggs. Eggs are cylindrical, 2.6 x 0.97 mm. Larvae are pelagic and as small as 4.1 mm standard length. Relative fecundity in the Gorgan Bay study in Iran was 290-550, mean 395.5 eggs (Afraei et al., 2001).
There is a pre-spawning migration into shallow water in April in the Caspian Sea. The male approaches the female from below, touching her lower jaw with his snout and leading her to a reed nest. Such behaviour occupies several hours to more than a day. The female enters the reed nest, inverts and deposits eggs on the roof of the nest. The underside of bivalve shells or gravel may also be used. The male then fertilises the eggs, inverting to do so. The eggs are laid in a line and the pair will form other lines. Egg patches may eventually contain as many as 3000 eggs at different developmental stages, which indicates males may spawn with several females. Spawning takes about 1.5 hours and the female gradually loses the brightness to her breeding colours and almost all breeding pigment is lost within a day. The pair mated 4 times over a period of 35 days (28 March, 6 April, 14 April and 2 May in an aquarium with fish from Lake Trichonis, Greece). The male remains mostly inside the reed nest, aerating the eggs and defending them against other gobies. At irregular intervals (8-60 minutes), the male would leave the nest for 1-5 minutes, occasionally feeding but always remaining near the nest.
Spawning in the pre-estuary area of the Volga River in the Caspian Sea takes place from the middle of April to the end of May, rarely to June. Spawning in the Caspian takes place in shallow water with some current at a depth of 0.15 to 1.5 m, a temperature of 15-27°C and over a wide range of salinities (Kevrekedis et al., 1990).
Parasites and predators
None reported for Iran.
Economic importance
None.
Conservation
This species seems to be rare in Iran with only Derzhavin's (1934) report and one other record. Its status cannot be assessed until more is known about its distribution. Kiabi et al. (1999) consider this species to be data deficient in the south Caspian Sea basin according to IUCN criteria. Criteria include abundant in numbers, habitat destruction, limited range (less than 25% of water bodies), absent in other water bodies in Iran, absent outside the Caspian Sea basin (sic). Vulnerable in Turkey (Fricke et al., 2007).
Further work
Further collecting will need to be carried out to determine its abundance and distribution in Iranian waters. Its small size may have led to it being ignored or confused with other gobies as young and it may be more widely distributed than current knowledge indicates.
Sources
Iranian material: Uncatalogued, 4, 22.2-35.3 mm standard length, Mazandaran, Gorgan Bay (no other locality data).
Knipowitschia iljini
Berg, 1931

Common names
gavmahi-ye Iljin, gel-ye mahi.
[Iljin's bychok in Russian, Iljin xulu in Azerbaijan].
Systematics
At least 4 syntypes are in the Zoological Institute, St. Petersburg (ZISP 22052) along with ZISP 24370 and possibly ZISP 24424 (Miller and Pinchuk in Miller, 2004). Although Berg (1931b) states "many specimens caught" this may refer to abundance while only 4 fish are illustrated and may be the type series. This species was described from "in the middle part of the Caspian Sea".
Key characters
The anterior oculoscapular lateral line head canals are more or less separate in the midline of the posterior interorbit, with double or separate pores κ, and canals extend anteriorly through interorbit of variable extent. The preopercular canal is absent or present (
Miller in Miller, 2004).Morphology
The first dorsal fin has 6-8 spines, usually 7, the second dorsal fin has 1 spine followed by 8-10 soft rays, the anal fin has 1 spine and 7-10, usually 9, soft rays, the pectoral fin has 15-18 rays. Lateral series scales 28-38. Vertebrae 31-33. Scales are ctenoid. The belly, head and the dorsal surface back to the second dorsal fin are all naked. There is no small, straight canal extending posteriorly from the junction of the orbital canals. The caudal fin is rounded and symmetrical. The anterior oculoscapular lateral line head canals do not unite in the posterior interorbit. Canals anterior to pores κ are typically absent and pore κ is double. The posterior oculoscapular canal is absent, the preopercular canal absent or present. Pore "a" is single or double with sometimes an additional pore or two on horizontal anterior oculoscapular canal behind eye.
Sexual dimorphism
Males have darker overall colouration and darker fins than the females (see below). The first dorsal fin in males has free tips and usually a dark spot at the rear margin. Males have a pointed genital papilla while females have a distally bifurcated papilla. The pelvic fins reach the genital papilla in males while in females they do not reach the vent. Females have a more slender caudal peduncle and the snout is more produced than in males.
Colour
The body overall has a brownish reticulate pattern. The back and flanks bear about 6-10 dark bands in males while in females bands are faint and restricted to the posterior flank. The male has a darkly edged caudal fin and the pectoral fin is dark ventrally. The dorsal fins have dark stripes and the anal fin has a dark stripe along its margin in males while females lack stripes. Females bear small dark spots irregularly distributed on the back and anterior flanks.
Size
Attains about 4.7 cm for males and 5.0 cm for females. Ragimov (2003) gives a maximum length of 5.2 cm.
Distribution
Ragimov (1965) reports this species between Kultuk and Astara in Azerbaijan and generally in the central and southern Caspian Sea, and mapped around the coast of the whole Caspian by Miller and Pinchuk in Miller (2004).
Zoogeography
This species is part of a marine fauna in the eastern Mediterranean and the Black-Caspian-Aral seas basins and its relationships are outlined under the genus description above.
Habitat
Found benthically and in the water column with adults in shallow coastal areas at 20-70 m with juveniles offshore as deep as 500 m Not recorded from fresh water.
Age and growth
Life span is estimated at one year (Miller and Pinchuk in Miller, 2004).
Food
Only mysids have been recorded from 8 fish examined (Miller and Pinchuk in Miller, 2004).
Reproduction
Gonad maturity begins in late August to early September, many fish are ripe by the end of January and most by the end of February. Spawning occurs from early April to possibly as late as September with a peak in May-June (Miller and Pinchuk in Miller, 2004). Fecundity reaches 2240 eggs and egg diameter 0.8 mm (Miller and Pinchuk in Miller, 2004).
Parasites and predators
None reported from Iran but sturgeon and Sander species eat this fish (Miller and Pinchuk in Miller, 2004).
Economic importance
None.
Conservation
Apparently rare in Iran but this may reflect collecting effort for this small species. Insufficient work has been devoted to this species to assess its status.
Further work
Further collecting will need to be carried out to determine its abundance and distribution in Iranian waters. Its small size may have led to it being ignored or confused with other gobies as young and it may be more widely distributed than current knowledge indicates.
Sources
Iranian material: None.
Knipowitschia longecaudata
(Kessler, 1877)
Black and Caspian seas, described from the southern and middle Caspian Sea, but no Iranian record. Ragimov (1965) reports this species between Kultuk and Astara in Azerbaijan. Sometimes spelt longicaudata erroneously in the literature.
This is an endemic Ponto-Caspian genus with 2-3 species, with one species being of uncertain status (see below). It is distinguished by having three (rather than the more usual two) transverse head papillae rows between rows b and d. Other characters include suborbital papillae in transverse rows, no row a, snout with longitudinal rows s1 and s2, no perianal organ; row 5i not below level of row 6i, 6i at or opposite end of row d, scales normal, anterior nostril elongate but not overhanging lip, and pelvic fins without lobules. A detailed, recent description is given by Miller in Miller (2004).
Mesogobius nigronotatus
(Kessler, 1877)
Caspian Sea basin, described from Fort Aleksandrov (= Shevchenko) on the Caspian Sea coast of Kazakhstan, and from the south Caspian Sea by Naseka and Bogutskaya (2009), but no Iranian record. The only specimen was lost (Reshetnikov et al., 1997). M. nonultimus may be a synonym (Pinchuk and Miller in Miller, 2004).
Mesogobius nonultimus
(Iljin, 1936)


Anzali Shore, January 2005, courtesy K. Abbasi

Anzali Shore, January 2005, courtesy K. Abbasi
Common names
[bychok neposledniy or seriy bychok-martovic in Russian; Caspian toad goby]
Systematics
Described from 24 miles southwest of Ulsky Bank, Caspian Sea, Turkmenistan. The type is lost. May be a subspecies of Mesogobius batrachocephalus (Pallas, 1814) although Miller revises his opinion in Pinchuk and Miller in Miller (2004) and recognises it as a distinct species. See also under M. nigronotatus above.
Key characters
It is distinguished by having three (rather than the more usual two) transverse head papillae rows between rows b and d. Nape scales weakly ctenoid, almost reaching median pore κ, snout less than twice eye diameter, lower jaw not protruding, and males with distinct breeding colours.
Morphology
First dorsal fin rays 6-7, second dorsal fin with 1 spine and 16-19 (usually 17-18) soft rays, and anal fin with 1 spine and 15-18 soft rays. Scales ctenoid except for a few, small cycloid scales on the opercle. The breast is naked. Lateral line scales 73-83. Lateral line system with 8 transverse suborbital neuromast rows. Three rows of free neuromasts on the opercle. The height of the second dorsal fin diminishes posteriorly. Head width 1.25 depth.
Meristics for an Iranian specimen: first dorsal fin with 6 spines, second dorsal fin 1 spine and 19 soft rays, anal fin with 17 soft rays and pectoral fin with 16 branched rays. Lateral series scales 80 and transverse scale rows 20.
Sexual dimorphism
Male spawning colour is distinctive and males are larger than females.
Colour
Overall colour is pale grey with dark bands, up to 6 on the body. The first band is in front of or below the first dorsal fin and is visible from above. Intensity of bands varies with ones below the middle of the second dorsal fin and before the caudal fin origin being the most intense. The first dorsal fin has two bands, the dorsal, pectoral and caudal fins are dark while the pelvic and anal fins are pale. Males have a dark, bluish-black spawning colouration with spots and bands on the upper body becoming indistinct.
Size
Reaches 17.4 cm.
Distribution
Probably the whole Caspian Sea basin. Known from the Turkmenistan and Daghestan coasts and from recorded between Kultuk and Astara in Azerbaijan (Ragimov, 1965). Pinchuk and Miller in Miller (2004) map this species from the entire Caspian Sea including the Iranian shore but only recently has it been recorded from Iran (H. Ahnelt, et al., 2007). K. Abbasi records it from the Anzali Shore (see photograph above).
Zoogeography
This species is related to Neogobius and Ponticola, more speciose genera of Ponto-Caspian gobies.
Habitat
This species has been collected down to 50 m, perhaps deeper, but little else is known of its habits. It is suspected to feed, winter and spawn in deeper waters without an onshore migration.
Age and growth
Maturity is attained in the second year of life.
Food
Unknown.
Reproduction
Fecundity reaches 1544 eggs, probably laid in a single batch. The spawning season off the Lenkoran coast is late March and early April, the earliest of all Caspian Sea gobies.
Parasites and predators
Unknown.
Economic importance
None.
Conservation
Numbers in Iranian waters and threats to this species need to be assessed.
Further work
The biology and distribution of this species in Iranian waters needs to be studied.
Sources
Based in part on Pinchuk and Miller in Miller (2004).
Iranian material: CMNFI 2006-0022, 1, 114.2 mm standard length, Gilan, southwest Caspian Sea (38º00'N, 49º30'E to 38º20'N, 50º00'E) (Ahnelt et al., 2007).
This genus of gobies is found in the Black and Caspian seas where there are about 14 species, some large enough to be the
object of commercial fisheries. The general Farsi name for fishes in this genus
is گاوماهي (gav mahi), سگ ماهي (sag mahi) or gel-ye mahi, not repeated under each species.
Berg (1948-1949) and various papers by Pinchuk (1976, 1977, 1991) recognise Neogobius as a genus but Vasil'yeva (1989) considers that the separation of Neogobius, even as a subgenus, is
doubtful and refers the species to Gobius. Simonovic (1999) briefly reviews the literature on Neogobius and its distinction from Gobius. Presence or absence of a swimbladder, egg size,
and presence or absence of pelagic larvae are characters which have been used to recognise genera or subgenera but these may be phylogenetically independent (Svetovidov, 1964b). Various subgenera may be
recognised within Gobius such as Gobius Linnaeus, 1758, Neogobius
Iljin, 1927 (type species N. fluviatilis), Apollonia Iljin, 1927 (monotypic for N. melanostomus
according to Pinchuk (1991) although Vasil'yeva (1989) and Dobrovolov et al. in Charlebois et al. (1997) add the species fluviatilis on the basis of osteology and electrophoretic
studies; and then later she (Vasil'eva, 1996a) places both these species in the subgenus Neogobius, again on osteological grounds), Ponticola
Iljin, 1927 including such species as
ratan (type species), and Eichwaldia for caspius (type species)(Eichwaldia is preoccupied by Eichwaldia Billings 1858 in fossil brachiopods, replaced by and objective
synonym of Eichwaldiella Whitley, 1930). Miller and Vasil'eva in
Miller (2003 - Vasil'eva's assignments) give the following subgenera, with definitions (these subgenera may eventually be elevated to genus once a thorough analysis is complete (see below); note
assignments conflict in part with the above:- Neogobius (with one species, caspius), Apollonia (fluviatilis and melanostomus), Babka (gymnotrachelus, not
in the Caspian Sea), Ponticola (the remainder - including species not in the Caspian Sea). Miller in Miller (2004) place Neogobius bathybius in the genus Chasar and this is followed
here. Neilsen and Stepien (2006) and Stepien and Tumeo (2006) used mtDNA cytochrome b sequence data and consider Apollonia (N. melanostomus and N. fluviatilis) to be highly
divergent from all other neogobiins, Mesogobius is the sister genus of Proterorhinus and a clade of these two genera is sister to Neogobius.
Neilsen and Stepien (2009a) using two mitochondrial and two nuclear genes
conclude that Neogobius, or in a restricted form Apollonia,
consists of fluviatilis, melanostomus and caspius, Babka
is monotypic (species gymnotrachelus), and Ponticola includes
cyrius, gorlap, ratan and syrman among others.
Mesogobius and Proterorhinus are sister groups. Tribes are also
delineated. Apollonia is synonymised with Neogobius.
Pinchuk (1991) states that many of the characters used to define genera are composed for convenience in identification rather than indicating natural relationships, i.e. the characters are simply those
used in keys. Ahnelt and Holčík (1996) call the taxonomy of this genus "variable, unstable and frequently uncertain" since variation between Black and Caspian Sea populations is usually very
small and since there are migrating and resident marine and freshwater populations of the same species. Miller and Vasil'eva in Miller (2003) state that the genus is paraphyletic (included species do not
share an autapomorphy) but retain it for convenience on morphological grounds, a practice followed here until a cladistic revision is performed.
These gobies are characterised by having an elongate body, compressed posteriorly, mouth of moderate size, 6 first dorsal fin rays, more than 10 rays in the second dorsal fin (large number of rays
in the unpaired fins according to Pinchuk (1991) compared to Gobius sensu stricto), anterior nostrils tubular but not very elongate, posterior nostrils near eye, ctenoid scales of moderate
size, a scaled nape and predorsal area, cheek naked, the pelvic fin anterior membrane may have obvious lateral lobes, no barbels, upper rays of pectoral fin may be free and filament-like (Pinchuk (1991)
states that they are always within the fin membrane or joined), teeth conical and in a few series, back muscles extend forward almost to the eyes, tongue not notched or only slightly notched, no
swimbladder in adults, 32-35 vertebrae modally (Gobius has only 28 and Proterorhinus 30-32 (Simonovič et al. (1996)), dorsal pterygiophore formula (3)22110, anal pterygiophores
before first caudal haemal spine 2 or 3, large, oligoplasmatic roe, and no pelagic larvae. Miller and Vasil'eva in Miller (2003) give details of neuromast organs as follows:- infraorbital neuromasts
typically in 7 transverse rows, 4 before and 3 above hyomandibular row b, and no row a. Row 7 is made up of a few to several papillae in a row descending postero-ventrally from just in front
of the anterior oculoscapular pore α. Rows 5i and 6i are separated with 5i well behind the anterior end of hyomandibular row b and with 6i reaching or falling short
of row b. Dorsal supraorbital rows o are separate in the dorsal midline. Hyomandibular row z ends near pore χ. Supratemporal accessory line x1
ending anteriorly behind pore β. Anterior oculoscapuler, posterior oculoscapular, and preopercular canals present. These have the pores σ,λ,κ,ω,α,β and ρ; θ
and τ; and χ, δ and ε. Anterior oculoscapular pore ρ and posterior oculoscapular pore θ well separated.
Neogobius is distinguished from the closely related genus Mesogobius Bleeker, 1874 by having a smaller number of infraorbital transverse rows of pit organs and the transverse parietal
posterior rows of pit organs are separated by a wide interspace (Pinchuk, 1991).
Simonovič et al. (1996) consider this genus of gobies to be a young one as evidenced by the distribution of species in the Caspian and Aral seas which closed recently. Simonovic (1999)
concludes that "Neogobius" (and Proterorhinus) evolved from a common ancestor in the Dacian Basin of the eastern Paratethys (now the Black Sea) or its tributaries during the early to
mid-Pliocene, as evidenced by their absence from Adriatic, Ionian and Aegean basins. Divergence probably occurred in freshwater, riverine habitats as development is direct without a pelagic larval stage
as found in marine gobiids. The hypersalinity of the isolated Black Sea in the late Miocene to early Pliocene was the causal event that led to rapid speciation of
"Neogobius" and the genus retained
direct development when it returned to the Black and Caspian seas. Dillon and Stepien (2001) using mitochondrial DNA suggest that the genera
"Neogobius" and Proterorhinus diverged about
5.2±1.0 MYA in the late Miocene/early Pliocene from a common ancestor shared with Gobius during the isolation of the Paratethys Basin from the Mediterranean Tethys Sea. This contradicts
suggestions that the neogobiins diverged from Gobius during the Quaternary interglacial connections of the Black and Mediterranean seas.
Masoumian and Aghl Mandi (2009) record Neogobius spp (presumably
including Ponticola spp.) infected with Ceratomyxa caspia, a
myxosporean, from the Iranian shore of the Caspian Sea. Neogobius caspius
Common names
gel-ye mahi Khazari (= Caspian goby), gavmahi-ye Khazari, gavmahi-ye Daryaye-e Khazar.
[xval xulu in Azerbaijan; Khvalynskii bychok or Hyrcanian goby in Russian; Caspian goby].
Systematics
No major synonyms and no types known. Described in Latin from "Hab. in Caspio mari, in sinu Bacuensi" (in the Caspian Sea, in Baku Bay, Azerbaijan).
Ghotb Razmjoue et al. (2008) examined the population genetic structure of
this species and found no polymorphism and no differentiation in the south
Caspian Sea population. Key characters
This species is separated from other Caspian gobies in Iran by the rear nostril being slit-like and far from the eye margin. It has a comparatively small mouth.
Morphology
Head canals and free neuromasts on the body are typical of a Neogobius. First dorsal fin spines 5-7, usually 6, second dorsal fin with 1 spine followed by 14-17 soft rays. The first ray of the
second dorsal fin may be elongated and branched at its tip and in such cases was counted as a "spine". Anal fin with 1-2 spines, usually 1, and 10-14 soft rays. Lateral series scales 57-71.
Predorsal area and nape scaled, anteriorly reaching to the orbit (postorbital section of the supraorbital canal of the cephalic lateral line system). Anteriormost scales cycloid. Scales have a
diamond-shaped pattern on the flank. Individual scales are squarish with the upper and lower margins straight to slightly rounded or, more rarely, each scale has an overall rounded shape. The posterior
margin has two straight edges meeting at the posterior centre. The posterior margin bears ctenii. The anterior margin is wavy where the radii terminate at indentations and there is a central protuberance.
Moderately numerous radii radiate anteriorly from the central point of the posterior margin, occupying the entire anterior field and spilling into the lateral fields. Circuli are fine. There is a dermal
papilla on the posterior edge of the gill chamber, just under the gill cover. The gill rakers are short anteriorly and posteriorly but vary in length and the longer central rakers reach the second adjacent
raker when appressed. Rakers are commonly forked or even antler-like. Dorsally the smallest rakers are difficult to distinguish from mouth cavity tubercles and only definite rakers were included in the
count. The pelvic fin anterior membrane has pointed lobes. The gut is an elongate s-shape with convolutions or twists along its length. The genital papilla is bifurcated at the tip.
Meristics for Iranian specimens: first dorsal fin spines 5(1), 6(37) or 7(2); second dorsal fin soft rays 14(2), 15(10), 16(27) or 17(1); anal fin soft rays 11(2), 12(25) or 13(13); pectoral fin rays
17(6), 18(29) or 19(5); lateral series scales 60(2), 61(4), 62(6), 63(8), 64(10), 65(6), 66(1), 67(1), 68(1) or 70(1); total gill rakers 8(1), 9(3), 10(17), 11(14), 12(4) or 13(1); and total vertebrae ?.
Sexual dimorphism
Colour differences are given below.
Colour
Overall colour is a dark grey or grey-brown with a clearly contrasting white to pearly white belly and lower head surface. The back may have 7-8 dark brown or grey saddles. The flanks have 6-10
obscure dark spots. The first dorsal fin has 2-4 dark brown stripes and a black spot over the last three rays. The black spot may not be very evident and merges with the general background pigmentation.
There are no clearly defined spots on the fins although the rays are brown in all but the pelvic and anal fins. The anal and pelvic fins are pearly-white. The caudal fin has a narrow whitish transparent
edge. The iris has a narrow golden ring. The genital papilla is white or grey. The peritoneum is silvery-brown in preserved specimens.
Breeding males are black all over, often including the margin of the first dorsal fin. First dorsal fin ray tips may be yellowish. The belly is whitish to pale grey in front of the pelvic fins. There
is a dark spot at the end of the first dorsal fin. The second dorsal fin has a narrow colourless margin but fin tips are not extended into free ends as obviously as in some Neogobius species. All
fins have a fringe effect although this is not well developed in the first dorsal fin. The caudal fin has a colourless or whitish margin. The dorsal fin spot is lost temporarily after spawning.
Size
Reaches 20.2 cm.
Distribution
This species is found in the Caspian Sea and is recorded between Kultuk and Astara in Azerbaijan (Ragimov, 1965). In Iran records are from both the southwestern,
south-central and southeastern Caspian Sea, the Anzali Shore and Gorgan Bay (Abdoli
and Naderi, 2009; K. Abbasi - see photograph above).
Zoogeography
Endemic to the Caspian Sea. Habitat
Found throughout the Caspian Sea but does not enter fresh water. It is found on sand or pebble bottoms, less often on stones and rarely on soft mud. They tend to remain inshore until the winter when
temperatures fall although high summer temperatures may drive them into deeper and cooler water for a short period (Pinchuk and Miller in Miller, 2003).
Age and growth
Sarpanah Sarkohi et al. (2010) found fish to reach age 5 years in the
Iranian Caspian Sea sample sites listed below. Food
Gut contents include polychaetes, crustaceans, small bivalve molluscs and fish. In some areas molluscs predominate, in others crustaceans.
The study by Sarpanah Sarkohi et al. (2010) of fish taken in bottom
trawlers from three stations at Astara, Anzali and Chaboksar found a relative
gut length of less than one suggesting this fish is carnivorous. Intensity of
fullness of guts was highest from April to October. The main food items were
molluscs (principally Cardium, Didacna, Mytilaster, Abra),
worms (principally Nereis, oligochaetes) and crustaceans (principally
Niphargoides, Petrcuma (sic), Paramysis) with food
preferences of 100%, 89% and 74% respectively. Some chironomids and gobiids were
also eaten. Reproduction
Egg sizes in Iranian specimens reach 2.1 mm and fish taken on 14 May have eggs 1.9 mm in diameter suggesting a spring to summer spawning season. This species approaches coastal areas in the southern
Caspian Sea at the end of March and the beginning of April, later than P. syrman and
P. gorlap. Females may have up to 1007 large eggs and up to 1985 small eggs, indicating
repeat spawning (Pinchuk and Miller in Miller, 2003). Sarpanah Sourkouhi et
al. (2008) examined fish form the Gilan coast and found the highest
gonadosomatic indices in June, suggested spawning occurs several times, found a
sex ratio of 1.47:1 in favour of females, an absolute fecundity of 212-1234
eggs, a relative fecundity of 18.73-29.89 eggs/g body weight and an egg diameter
range of 0.06-0.23 mm. Parasites and predators
Sattari et al. (2002) and Sattari (2004) records the presence of the nematode, Eustrongylides excisus, in this species. This parasite can damage muscles in commercial species and render them unsuitable for sale. This
species is eaten by the Caspian seal and by sturgeons. Daghigh Roohi and Sattari
(2004) record Eustrongylides excisus, and Dichelyne minutus from
this species in the southwestern Caspian Sea of Iran. Sattari et al. (2004; 2005) surveyed this species in the inshore area of the Caspian Sea, recording Eustrongyloides excisus and Dichelyne
minutus.
Economic importance
Not reported as of economic importance in Iran, it has been caught commercially in the former Soviet Union and on rod and line. It does form part of the diet of sturgeons (Pinchuk and Miller in Miller,
2003; Ghotb Razmjoue et al., 2008).
Conservation
Trends in numbers of individuals of this species and threats to it have not been examined so its conservation status is unknown.
Further work
The biology of this species needs investigation.
Sources
Pinchuk and Miller in Miller (2003) is the most recent summary on this species.
Iranian material: CMNFI 2006-0022, 1,124.8 mm standard length, Gilan, south-west Caspian Sea off Astara (38°00’N, 49°30’E to 38°20’N, 50°00'E);
CMNFI 2006-0027, 2, 113.4-121.1 mm standard length, Gilan, Talesh (37°48’N, 48°55’E);
NMW 80605, 1, 116.1 mm standard length, Mazandaran, Now Shahr beach (36°39’N, 51°31’E);
NMW 95073, 1, 92.8 mm standard length, Mazandaran, Gomishan lagoon near estuary of Gorgan River (36°59’N, 54°00’E).
Neogobius melanostomus
Common names
gavmahi-ye gerd (= round goby), gel-ye mahi gerd
(= round goby), gavmahi domgerd, gavmahi-e-domguerd, sebele.
[xazar kirda xulu in Azerbaijan; Kaspiiskii bychok-kruglyak or Caspian round goby in Russian; black spotted goby].
Systematics
Gobius melanostomus was originally described from Sevastopol and Balaklava, Ukraine, on the Black Sea. No types known.
Gobius affinis Eichwald, 1831 described in Latin from "Hab. in Caspii maris sinu bacuensi, balchanensi" (Baku and Balkhan bays in the Caspian Sea, no types known) and Gobius
sulcatus Eichwald, 1831 described in Latin from "Hab. in Caspii maris sinu balchanensi" (Balkhan Bay in the Caspian Sea, no types known), are synonyms. The Caspian Sea
population was referred to Neogobius melanostomus affinis (Eichwald, 1831) but subspecies are no longer recognised (Iljin, 1956; Pinchuk, 1976).
However, Brown and Stepien (2008) conclude on DNA evidence that Black and
Caspian Sea populations diverged ca. 350,000 years ago and this supports
subspecies separation. Key characters
Pelvic fin anterior membrane with rounded and shallow lateral lobes; lobes not more than one-sixth width of anterior edge of membrane, or lacking entirely; nape scaled and at least anterior nape scales
cycloid; first dorsal fin with large dark spot at rear; and lateral series scales usually 49-55.
Morphology
This species is separated from other Caspian gobies in Iran by having a small mouth, rounded, shallow, poorly-expressed lobes to the pelvic anterior membrane, scales in lateral series 41-59 (usually
47-55), generally at the low end of this range, mid- and anterior nape scales are cycloid as are most of those on the gill cover, throat, pectoral in bases and part of the abdomen, the pelvic disc is
0.6-0.8 pelvic base to anal fin length, the jaw angle is below the level of the anterior part of the eye, the posterior nostril is close to the eye margin, and the first dorsal fin
has an obvious dark spot posteriorly.
First dorsal fin spines 5-8, usually 6, second dorsal fin with 1 spine and 11-18 soft rays, anal fin with 1 spine and 9-14 soft rays, and pectoral fin with 16-20 branched rays. Scales appear as
diamond-shaped on the flank since the posterior margin has two straight edges coming to a central point. However scales can be quite rounded even adjacent to those forming a diamond pattern. The
posterior margin bears ctenii. Dorsal and ventral margins are rounded and the anterior margin is indented above and below a central protuberance. Radii radiate anteriorly from the centre of the
posterior margin. Circuli are fine. Gill rakers 9-13, presumably on lower arch only, see below. Gill rakers may reach the second adjacent raker when appressed and although usually
regular can be branched and variable in size along the arch. Vertebrae 31-34.
Head depth at the eyes is 0.9-1.2 head width between the upper origins of the opercles. The interorbital width is 0.8 to slightly less than eye diameter. The snout is longer than the eye, 1.1-1.4
times. The upper lip narrow slightly rearward and is about half the lateral preorbital area (between lip and eye). Caudal peduncle depth is two-thirds its length. The tongue is truncate or slightly
notched. The posterior teeth on the dentary bone are smaller than the anterior teeth and all are blunt. Pharyngeal teeth are molariform, adapted to crushing molluscs. The anal papilla is bifurcated at
the tip. The gut is short and roughly s-shaped with some convolutions, particularly on the posterior section.
The chromosome number for "N. m. affinis" is 2n=46 with 46 chromosomal arms (Ráb, 1985; Vasil'ev and Grigoryan, 1993; Simonovic, 1999).
Meristics for Iranian fish:- first dorsal fin spines 5(6) or 6(34); second dorsal fin soft rays 12(1), 13(2), 14(18), 15(18) or 16(1); anal fin soft rays 10(3), 11(17), 12(19) or 13(1); total pectoral
fin rays 17(11), 18(28) or 19(1); lateral series scales 48(1), 49(3), 50(8), 51(4), 52(8), 53(4), 54(5), 55(4), 56(2) or 57(1); total gill rakers 11(5), 12(15), 13(11), 14(7) or 15(2); total vertebrae ?.
Sexual dimorphism
Females are smaller than the male. Mature males have larger dorsal, pectoral and anal fins, swollen cheeks, and colour is different (see below).
Colour
Overall colour is yellowish-grey or brownish-grey (fawn) with 8-9 large, dark brown, blue-grey to black flank blotches. There are 4 saddles across the back. The first dorsal fin has a black blotch
posteriorly over the last two rays (and sometimes on the interradial membrane between rays 4 and 5), and the blotch or spot may have a white border in juveniles. The spot can be absent as reported by
Jude (1997) for fish introduced to the North American Lake Erie. The sides of the head have oblique, curved bands or a reticulate pattern of a rusty to gingery tint on a bluish-grey background. The
pattern is darker on the opercle and snout. The first dorsal fin may be tinged green and has 3 rusty-brown bands and a narrow margin may be a rusty or yellowish colour. The second dorsal and caudal fins
may have a light margin even in non-spawning individuals. The second dorsal fin has 3-4 rows of small rusty spots. The caudal fin also bears rusty spots anteriorly but not in very definite rows. The anal
fin is grey to dark brown. The pectoral fin lacks spots and is almost colourless but may be tinged with yellow. The pelvic disc is grey with a rusty colour in the centre. The genital papilla is grey or
unpigmented. The iris is yellow to orange. The peritoneum is a silvery brown.
Colour varies with habitat. In bays with algal growth fish are darker, blue-grey tints are lost, there are more yellowish-brown colours and there is an increase in number and size of dark-brown spots.
Others may have an olive-green tint.
When spawning, males are charcoal black overall, sometimes with a bluish tint, and with indistinct yellow spots. The dorsal, caudal and anal fins are white-edged or yellow, although the first dorsal
margin can be black and there may be practically no margin to the anal fin. The black spot on the first dorsal fin can still be seen against the light. There are no free tips to fin rays according to
literature reports but Iranian specimens have all fin tips separated to form a fringe.
The young are a slate grey overall with a mottled and blotched flank. The mid-flank blotches may form a series of about 6. There is a spot at the upper base of the pectoral fin. The dorsal fin spot is
centred on the membrane between the last two rays but spills over onto the adjacent membranes, particularly the posterior one. Fins have pigment on the rays and membranes with no clear pattern. The dorsal
fins are darker than the anal which is darker than the pectoral or pelvic. There is a bar from the lower eye margin to the upper lip just anterior to the mouth corner.
Size
Reaches 29.0 cm total length. Freshwater fish are smaller than marine ones.
Distribution
Found from the Aegean to the Black and Caspian seas. Introduced to the Aral Sea in the 1950s but increasing salinity in the 1980s eliminated the population. Now in the Baltic Sea via rivers and canals
or in ship ballast. Also present by introduction in the Great Lakes of North America where first reported in 1990, apparently from several sources (Brown and Stepien, 2006).
Reported from a wide range of rivers along the Caspian coast of Iran
including the Gorgan, Tonekabon, Safid, Dyusan, Shimrud, Chemkhaleh, Rud-e Sere,
Kargan, Navarus, Chalus, Babol and Haraz rivers, the Anzali Mordab and the
Anzali Shore, Gorgan Bay, the southeast Caspian Sea, southwest Caspian Sea and south-central Caspian Sea (Holčík and Oláh,
1992; Abbasi et al., 1999; Kiabi et al., 1999; Abdoli, 2000; Jolodar and Abdoli, 2004;
Abdoli and Naderi, 2009; K. Abbasi - see photograph above). Apparently absent from the Atrak River in Iran
and of uncertain presence in the Aras River (Abdoli
and Naderi, 2009). Zoogeography
See genus account above.
Habitat
This species is found in inshore waters at depths to about 20 m, sometimes to 70 m or even 200 m in deeper sea areas in winter, on rock, gravel, shell, sand or silt bottoms. Aquarium specimens are seen
to hide in crevices and under rocks. In the Caspian Sea, it may be found among eelgrass, Zostera. It also enters rivers. Larvae live near the bottom. The species prefers littoral areas where wave
action keeps oxygen levels high and decaying material is reduced. It leads a sedentary life (Moskal'kova, 1996). It may be found from fresh waters to salinities of 40.5‰ from -1 to 32°C, and respiring
through the skin can tolerate low oxygen conditions from 0.3-0.9 ml/l (Moskal'kova, 1996; Jude, 1997).
In the North American Great Lakes basin, where it is an exotic, this species prefers rocky habitats and is more active during the day than the night. The mean density in one river was approximately 7
gobies/sq m and can reach as high as 90 gobies/sq m since they aggregate. This goby has a high site fidelity in mark-recapture studies (Ray, 1997; Ray and Corkum, 1997a).
The gobies can tolerate a flow of 0.34 m/s for 3-4 minutes but at higher levels they retreat to the bottom and brace themselves against the current using the pectoral fins.
The onshore spawning migration in the southern Caspian Sea occurs in spring, later than
P. syrman and P. gorlap. A spawning area is chosen on the basis of stones being present rather than
depth. Larger males approach the shore first. There is an offshore movement as winter begins and in the hottest part of summer.
Age and growth
Maturity is attained in the second year by females and in the third year by males but may be as early as 1 year. Life span can exceed 5 years for females. Males die after spawning. Faster growth
results in a shorter life span. At the end of the first year of life they can reach 5.5-6.0 cm (Moskal'kova, 1996). In North America, populations of this exotic are dominated by age one fish (MacInnis
and Corkum, 1997a). Growth is slower in North America and maximum size is less than in its native habitat, probably because temperatures are lower than in Europe (Jude, 1997).
Abdoli et al. (2006) examined fish from the Gomishan and Miankaleh
wetlands and found a length (L)-weight (W) relationship of W = 0.0091L3.17
for males, 0.0249L2.7 for females and 0.0112L3.08 for
sexes combined. Food
Abd Elahpour Biria et al. (2010) examined samples from 0-5 m, 5-10 m
and 10-15 m from Astara, Anzali and Chaboksar coasts. Molluscs and benthic
crustaceans were the preferred foods, feeding decreased with increased
temperature, there was no difference in feeding intensity between sexes but some
differences were noted based on station and sampling depths. Abdoli et al.
(2012) found fish in the Gomishan Wetland preferred bivalves, chironomids and
amphipods with bivalves dominating numerically, especially in summer, and
occurring in all seasons. Its feeding strategy was mixed in all seasons. Molluscs predominate with significant amounts of crustaceans in the north Caspian Sea but this can vary annually with crustaceans becoming predominant. They feed most heavily in the post-spawning
period in July and August (Opalatenko, 1979). Other important foods are bivalves, such as zebra mussels, snails, polychaetes, chironomids, other aquatic insects, goby eggs and small fishes. Attached
clams and mussels are bitten off the substrate by the large jaw teeth and crushed by the pharyngeal teeth (Moskal'ova, 1996). Molluscivory is also documented for Danube River populations (Simonovič
et al., 2001) and for Baltic Sea populations (Skora and Rzeznik, 2001) and molluscs appear to be the preferred diet wherever this species is found. An exception may be the southern Caspian Sea
where one study found crustaceans to predominate (Pinchuk et al. in Miller, 2003).
In the Caspian Sea, introduced species, such as crabs, the worm Nereis diversicolor and the mollusc Abra ovata, are also eaten (Kosarev and Yablonskaya, 1994). Iranian specimens contain
plant fragments, mollusc shells, shrimps, fish remains, aquatic insects and polychaetes.
In North America, immature round gobies are the most successful predators on round goby eggs. Round gobies are said to be aggressive and will take a baited hook. However this goby may need to be
stationary to detect prey. Nocturnal feeding occurs. The blunt teeth are indicative of a primarily mollusc diet.
Experimental studies in North America have shown this species to prefer individual and clumped zebra mussels to other foods such as sphaeriid clams. Feeding rates vary between 36 and 47 mussels
(4.5-12.5 mm long) per day for fish 6-10 cm long, over 100 mussels per day when the mussels are smaller than 4 mm (Ghedotti et al., 1995). Mussels can remain in the mouth and pharynx for less than
one hour to over 12 hours; shells are then split and ejected. However some are crushed and passed through the digestive tract. In the upper Detroit River, Canada, round gobies eat zebra mussels (58%),
snails (6%) and other invertebrates (36%) such as aquatic insects, softshelled crayfish and zooplankton. In laboratory studies, round gobies ate an average of 1 g of mussels in 24 hours, smaller mussels
being preferred (Ray and Corkum, 1997b). However, field studies in Lake Michigan show that round gobies are unlikely to remove zebra mussels (also a pest in North America) from a habitat (Djuricich and
Janssen, 2001). These gobies are also predators on lake trout eggs under laboratory conditions in North America (Chotkowski and Marsden, 1999). They are also reported to eat sturgeon eggs in North America.
Reproduction
There is a movement inshore to waters about 0.2-1.5 m deep for spawning, males preceding females. Spawning takes place at the end of April to September in near shore areas of the north Caspian Sea and
over a similar period in the Baltic Sea. Iranian specimens have well-developed testes on 10 April and spawning may occur earlier than in the North Caspian. There may be repeat spawning, up to 6 times
every 18-20 days, as indicated by captive specimens. Peak spawning occurs at 15°C (range 10-30°C). Eggs are attached to or under rocks, in cavities, under logs or in such objects as cans and are
aggressively guarded by the male. Early maturity, a very long spawning season and aggressive defense contribute to high egg, larvae and fry survival. The male prepares the nest site with a secretion from
a cement gland which is coated over the site. As the female lays each egg, it is glued to the underside of the nest cavity roof in single layer rows. Egg clutches can be transported on the hulls of ships
(Ahnelt et al., 1998). Fecundity reaches 6,177 eggs. Eggs are pale yellow, orange or pink, with red pigmentation. the eggs are ovoid with a sharp apex and measure 3.9 by 2.2 mm. Details of
development are given by Moskal'kova (1996).
In North America, the species has been shown to be a multiple spawner with an extended reproductive season and male guarding of eggs (MacInnes, 1997). Here maturity occurs as early as year one and 43
mm standard length, a year earlier than in their natural habitat. Spawning extends from May to early August. Mean fecundity is 198 eggs. Each nest is utilised by large numbers of females since as many as
an estimated 9462 eggs were found in each artificial nest site (MacInnis and Corkum, 1997b). Such a nest site can lose 50-70% of the eggs to predators while smaller clutches lose few eggs.
Jude (1997) notes that in North America some males do not defend a nest, the reason being unknown. This may well be due to such factors as availability of suitable spawning sites, large size or
aggressiveness. He reports that after reaching a large size, males spawn once and then die, although females spawn up to six times through spring and summer, about every 20 days. Elsewhere males guard
the nest from predators and fan the eggs to oxygenate them and to reduce siltation and fungal infection. Males will eat infected and unfertilised eggs. Males defend the nest by flaring the gills,
spitting sand, darting at nearby intruders, biting, and growling for 1.0-1.5 seconds loud enough to be heard 10 m away. Nest sites in North America may be as deep as 7 m (Corkum and Wickett, 1998).
Males make several sounds including one attracting females to nest sites and another to intimidate other males. Females respond to male calls by a quieter sound (Jude, 1997).
Parasites and predators
Fil mahi (Huso huso), chalbash (Acipenser gueldenstaedtii), uzun burun (Acipenser stellatus), mash mahi (Aspius aspius), ordak mahi (Esox lucius), suf
(Sander lucioperca) and seals are predators. Daghigh Roohi and Sattari
(2004) record Dichelyne minutus and Corynosoma strumosum from this
species in the southwestern Caspian Sea of Iran. Sattari et al. (2004; 2005) surveyed this species in the inshore area of the Caspian Sea, recording Dichelyne minutus.
Sattari et al. (2007) report the nematode Eustrongylides excisus
for this species in Iranian waters. Economic importance
Robins et al. (1991) list this species as important to North Americans. Importance is based on its use in aquaria, as food and because it has been introduced outside its natural range. It is
caught by anglers in some parts of the Caspian Sea basin.
It is present in the North American Great Lakes and is expanding (Ray and Corkum, 2001; Clapp et al., 2001; Jude et al., 1992; Jude, 2001). It is an introduction from the
ballast waters of ships from multiple sources, perhaps including the Black Sea (Dillon and Stepien, 2001). Round gobies may displace native fish, compete for food and eat their eggs and young, and are the
target of major efforts to study and contain them (Arrigoni and Berg, 2001; Charlebois et al., 2001; Diers et al., 2001; French and Jude, 2001; Janssen and Jude, 2001; Kaur et al.,
2001; Schreier et al., 2001; Wolfe, 2001). They may also accumulate and pass on large concentrations of such chemicals as PCBs to sport fishes in North America (Jude, 1997). An extensive literature
is now available on the Great Lakes populations.
A summary of invasions and biology in North America and Europe is given in
Kornis et al. (2012). It is an important food species in the Black Sea marketed fresh and formerly canned in tomato sauce. In the Sea of Azov in 1950-1960, gobies made up the bulk in local fisheries and round gobies
comprised almost 90% of a catch exceeding 90,000 t (Moskal'kova, 1996). Freyhof
(2011) records that this species is salted, dried and consumed with beer in the
Azov Sea area. Commercial catches in the Caspian Sea have comprised up to 88.2% of goby catches but pollution has severely depleted stocks
(Pinchuk et al., 2003).
Conservation
Kiabi et al. (1999) consider this species to be of least concern in the south Caspian Sea basin according to IUCN criteria. Criteria include medium numbers, habitat destruction, widespread range
(75% of water bodies), absent in other water bodies in Iran, and absent outside the Caspian Sea basin (as N. m. affinis).
Further work
Biology in Iran has not been studied as well as elsewhere.
Sources
Charlebois et al. (1997) and Pinchuk et al. (2003) give general reviews of biology and are excerpted briefly above.
Iranian material:
Neogobius pallasi
Common names
gavmahi-ye tireh (= dark goby), gel-ye mahi
rudkhanehai (= river goby), gavmahi-ye sheni, sebele.
[gumlur xulu in Azerbaijan; Kaspiiskii bychok-pesochnik or Caspian sand goby, rastrepka (= a person with untidy hair in reference to the fringed ends of the fin rays), chernysh (= darkie), khylak (when
emaciated), all in Russian; Caspian monkey goby].
Systematics
Gobius fluviatilis was originally described in part from near the mouths of rivers falling into the Black Sea and similarly the Caspian Sea. No types are known.
Neogobius fluviatilis pallasi (Berg, 1916) was the subspecies of the Caspian Sea basin. Kottelat and Freyhof (2007) recognise pallasi as the Caspian Sea species and restrict fluviatilis
to the Black Sea basin. The type locality is the Caspian Sea, mouths of the Ural and Volga and syntypes are in the Zoological Institute, St. Petersburg under ZISP 2195, 2204, 23137, 30729, 30736, 30738,
30919, 30920, 30924-26. 33182, 34276, 34277 (Eschmeyer's "Catalog of Fishes", downloaded 23 August 2007).
Abd Elahpour Biria et al. (2009) examined fish from 4 localities on
the Gilan coast using morphometric characters and found 3 different populations
in Astara, Anzali and Chamkhaleh and Chaboksar. Key characters
Pelvic fin anterior membrane with rounded and shallow lateral lobes. less than one-fifth rear edge width; pelvic fin disc 0.9 to more than distance to anal fin origin; nape with ctenoid scales; first
dorsal fin without large dark spot (except in juveniles); lateral series scales usually 55-70; pelvic fin membrane anterior lobes small, rounded; angle of jaw anterior to orbit; and two transverse
infraorbital papillae rows below longitudinal hyomandibular row b.
Morphology
The head width (between upper origins of opercles) is about equal to head depth (at eyes). The interorbital distance is not more than 0.75 eye diameter. The top of the head and the occiput are scaled
with ctenoid scales. The upper lip is expanded towards the mouth corners, sometimes narrowing at the very end. The angle of the jaws reaches back at a level between the eye and the posterior nostril. The
snout is longer than the eye diameter (1.6 times). The upper lip is not swollen at the angle, and measures 0.4-0.6 lateral preorbital area (between lip and eye). The first dorsal fin is high with an acute
anterior profile, the first branched ray about twice as long as the penultimate ray. The pelvic fin has small, rounded lateral lobes (< 0.2 rear edge width) and almost reaches the anal fin (0.9
distance) or extends beyond the anal fin origin. The caudal peduncle depth is 0.5-0.6 length.
First dorsal fin spines 5-7, usually 6, second dorsal fin with 1 spine followed by 11-18, usually 15-17, soft rays, anal fin with 1 spine and 11-17 soft rays, usually 13-15, and pectoral fin rays 16-19.
Lateral line scales 49-69, mostly 52-65. Gill rakers 6-10. Vertebrae 31-35. The chromosome number is 2n=46, with 46 acrocentric chromosomes and 46 chromosomal arms (Grigoryan and Vasil'ev, 1993; Klinkhardt
et al., 1995; Simonovic, 1999).
N. pallasi is distinguished by the dorsal fin spot in young fish and an average number of lateral series scales being 53 as opposed to 62 for Black Sea fish.
Sexual dimorphism
See under colour. Males become slimmer at spawning and their are morphometric differences between the sexes (Pinchuk et al. in Miller, 2003).
Colour
Overall colour is pale and sandy, in cantrast to other gobies. On the flanks and over the back there is a series of 8-9 brownish elongate spots or blotches. The belly, lower head and genital papilla
are pearly white. A dark bar runs from the eye antero-ventrally to the lip. The first dorsal fin has a dark spot posteriorly, although it may be absent in some adults, and is characteristic of the
former subspecies pallasi, and found in small fish in the southern Caspian, persisting in adults in the northern Caspian (Pinchuk et al. in Miller, 2003).
The first dorsal fin has 1-2 rows of grey to black spots and the second dorsal fin has 1-3 rows of pale grey spots. The caudal, anal, pectoral and pelvic fins are mostly grey on the rays and immaculate.
Breeding males become black or blue-black with the dorsal, caudal and anal fins bearing a yellow-orange margin. Some fish have a very narrow colourless margin to the pectoral and pelvic fins. The second
dorsal fin may only be yellowish on its first ray, the remainder being white and the caudal margin may be colourless. Rays in the second dorsal fin of breeding males become extended into free tips.
Size
Reaches 24 cm total length although in the Caspian the largest fish are 16.0
cm males (Pinchuk et al. in Miller, 2003).
Distribution
Found in the Caspian Sea and its basin. In Iran, it is found in rivers along the Caspian coast including some upper reaches, the
Aras River middle reaches and its tributary the Qareh Chai, the Anzali Mordab
and Shore, Gorgan Bay,
at Bandar Gaz, in the Gorgan, Madarso (or Madar Su), Tajan, Babol, Haraz, Sardab,
Tonekabon, Pol-e Rud and Safid rivers (including their middle to upper reaches
such as the lower Qezel Owzan and Shahrud of the Safid River basin), the southeast Caspian Sea, the southwest Caspian Sea and the south-central Caspian Sea (Shamsi et al., 1997; Abbasi et al., 1999; Kiabi et
al., 1994; 1999; Abdoli, 2000;
Abdoli and Naderi, 2009; K. Abbasi - see photograph above).
Note that Abdoli (2000) records it from the upper Atrak River in contrast to
earlier authors.
Zoogeography
The southern Caspian Sea and its tributary rivers is inhabited by a distinct subspecies, presumably on account of isolation from populations in the Black Sea. See genus account above.
Habitat
This species is found in inshore brackish waters and enters rivers. Salinities up to 46.9‰ are tolerated in marine bays. It is found down to 25 m although depths of 0.5-10 m are preferred, shallower
in spring (0.5-5.0 m), slightly deeper in summer and deeper still in fall (5-10 m)(Pinchuk et al. in Miller, 2003). It is found over sand, mud and sandy-shelly bottoms. In the Madarsoo stream of
Golestan National Park, abundance ranged from 0.1 to 0.75 fish per sq m and biomass from 0.21 to 4.88 g/m2 (Abdoli et al., 2002). Floods in this stream reduced the population, presumably
through destruction of the substratum and its food supply. Smaller fish were caught in riffles while larger fish lived close to to the river bank in depths up to 0.5 m. Abdoli and Rahmani (2001) found this
species to represent 95% of the goby catch in the Madarsoo stream, with N. melanostomus occupying the remainder.
Age and growth
Sexual maturity is attained at 2 years of age (9-12 cm for males, 8.5-10.5 cm for females) but may be as early as 1+ year (5.5 cm), particularly in the Caspian. Spawning males near Bandar-e Anzali were
12.4-15.2 cm long. Life span is 6 years but perhaps only 2+ years in the Caspian (Pinchuk et al. in Miller, 2003). Abdoli et al. (2002) found a sex ratio of 1 male to 3 females in the
Madarsoo stream. Patimar et al. (2008) reported 5 age groups in fish from
the Zarrin-Gol River in the eastern Alborz Mountains. Maximum total lengths were
12.1 cm in males and 10.8 cm in females, both 4+ years old. Fish aged 2+ and 3+
years were the most frequent in the population. sex ratio was equal and growth
was negative allometric. Condition factor was highest in early September. Males
showed a higher growth rate than females after age 1+ years. Abdoli et
al. (2006) examined fish from the Gomishan and Miankaleh wetlands and found
a length (L)-weight (W) relationship of W = 0.0059L3.291 for males,
0.0128L2.92 for females and 0.0057L3.29 for sexes
combined. Rahmani and Abdoli (2009) studied fish from the Madarsu Stream in
Golestan National Park. The population was dominated by males (75.3%),
weight-length relationships were W= 0.004L3.32 for males and W =
0.00L4.02 for females, showing positive allometric growth, the oldest
fish from otolith readings was 3+ years for both sexes with 2+ the most abundant
age, von Bertalanffy growth parameters were L(t) = 26.78(1-e-0.1(t+1.776)
for males and L(t) = 29.04(1-e-0.1(t+1.472) for
females Food
Diet items include crustaceans with lesser amounts of molluscs, polychaetes, chironomids and small gobies in the north Caspian Sea. They feed most heavily in the post-spawning period in July and August
(Opalatenko, 1979). The quantity of amphipods eaten increases with age while the amount of mysids decreases. Introduced species, such as crabs, the worm Nereis and the mollusc Abra, are also
eaten (Kosarev and Yablonskaya, 1994). In the Madarsoo stream of Golestan National Park in Iran, Ephemeroptera and Chironomidae are the most important food items but also included Trichoptera, other
aquatic and terrestrial insects, crustaceans (including freshwater crabs), arachnids, worms and snails (Abdoli and Rahmani, 1999; Abdoli and Rahmani, 2001; Abdoli et al., 2002).
Alavi Yeganeh and Kalbasi (2006) found fish in shallow water at Noor fed on
amphipods, fish larvae, barnacles, palaemonid shrimps, mytilid bivalves, xanthid
crabs, cardid bivalves, fish eggs and oligochaete worms, in order of preference.
Diet varied with season, males feeding less during spawning for example Patimar et al. (2008) found the most frequent food items of fish in the
Zarrin-Gol River were trichopterans, molluscs, chironomids and gammarids. Abdoli
et al. (2012) found fish in the Gomishan Wetland preferred chironomids,
bivalves and amphipods, most frequently chironomids, and showed a mixed feeding
strategy. Reproduction
Spawning can take place from late April to September, varying with locality. Peak spawning in the Volga River delta is in May. Spawning is recorded at 18-26ºC, 20-28‰ and 0.5-1.3 m (Pinchuk
et al. in Miller, 2003). In the Madarsoo stream of Iran, spawning peaked in March with an absolute fecundity of up to 532 eggs (Abdoli et al., 2002). Fecundity reaches 1025 eggs in the
southern Caspian (perhaps 12,800 eggs elsewhere) and egg diameter 2.1 mm. Yellow-amber or pale pink eggs are laid on or under stones and plants. Nests are excavated by the male under natural or artificial
objects. The male defends the nest and larger males are more successful. However, the stress associated with constant defense against predators and competing males leads to extreme emaciation. Nests
contain more eggs than fecundity would indicate suggesting that more than one female lays eggs in a nest (Pinchuk et al. in Miller, 2003).
Patimar et al. (2008) found the Zarrin-Gol River population to have an
average absolute and relative fecundity of 508.47 eggs and 61.27 eggs/g. The
reproductive season was March to April. Parasites and predators
Pazooki and Aghlmandi (1998) found the nematode Dichelyne minutus infecting the intestine of 6.6% of fish examined from the Tajan River in Mazandaran.
and Sattari et al. (2002) and Sattari (2004) records the presence of the
nematode, Eustrongylides excisus, in this species. This parasite can damage muscles in commercial species and render them unsuitable for sale. Fil mahi (Huso huso), chalbash (Acipenser
gueldenstaedtii) and suf (Sander lucioperca) are predators. Sattari et al. (2004; 2005) surveyed this species in the inshore area of the Caspian Sea, recording Eustrongyloides excisus
and Dichelyne minutus.
Daghigh Roohi and Sattari (2004) record Eustrongylides excisus,
Dichelyne minutus and Corynosoma strumosum from this species in the
southwestern Caspian Sea of Iran. Economic importance
This is the most economically important goby in the north Caspian Sea according to Berg (1948-1949).
Iljin (1956) considered this an error; it is the most numerous species but commercial fishing does
not occur there. Pinchuk et al. in Miller (2003) state however that it comprised 54% of the entire goby catch, although only 7.7% in southern Dagestan. It is food for sturgeons
and Sander lucioperca.
Conservation
Kiabi et al. (1999) consider this species to be of least concern in the south Caspian Sea basin according to IUCN criteria. Criteria include abundant in numbers, habitat destruction, widespread
range (75% of water bodies), absent in other water bodies in Iran, and absent outside the Caspian Sea basin (as N. f. pallasi).
Further work
On of the better known gobies in Iran but could still use work on its biology.
Sources
(Eichwald, 1831)



(Pallas, 1814)




(Berg, 1916)


Genus Periophthalmus
Bloch and Schneider, 1801
This mudskipper genus is found in the eastern Atlantic, Indian and western Pacific oceans. There are 12 species in the genus as yet undefined by a synapomorphy (Murdy, 1989). The pelvic fins are only partially united or may be totally separate. There are usually 12-14 pectoral fin rays, 4-17 first dorsal fin spines, and 8-13 second dorsal and anal fin rays. Scales are cycloid and cover the whole body and head except for the snout, isthmus and interorbital region in most species. Caninoid teeth are present in both jaws as a single row, with the anterior ones larger and pointed. The eyes are erectile and have a dermal cup covering the lower portion. There is a median fleshy ridge anterior to the eyes.
Mudskippers are found on muddy shores associated with mangroves but also venture into rivers. They spend much time out of the water.
Periophthalmus waltoni
Koumans, 1955
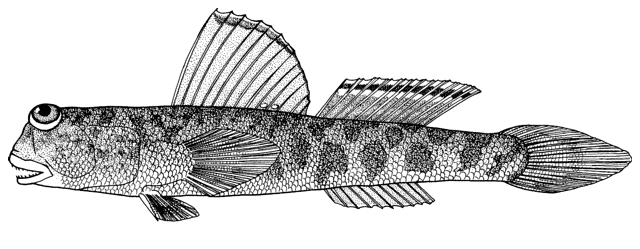
Common names
gel-cheragh (= mud-eater, mud-grazer), mahi-sag (= dog fish), gel khorak.
[shelambo, abou-shlembo or abu-shelamboo in Arabic; gullo in Pakistan; dark-blotched mudskipper, Walton's mudskipper, spotted mudskipper].
Systematics
This species was originally described from Iraq and Pakistan. The type locality is unclear as the information given by Koumans (1955) gives more than one locality, as follows:-
"Habitat.-Mesopotamia, Karachi.
I have seen 2♂ and 2♀. These were from the following localities:-
Indian Museum Collection.
Fao estuary of the Shatt-el-Arab, Mesopotamia, ♀ H. J. Walton.
Karachi ♂ and ♀
W. D. Cumming.
Mouth of Shatt-el-Arab ♂
W. T. Blanford.".
The type locality could be Fao and the holotype is the female specimen caught by H. J. Walton and named for him, the remainder being paratypes - by implication. Alternatively and probably correctly, these are all syntypes as no holotype was formally designated. The date of publication of Koumans work is 1955, not 1941 as cited by Murdy (1989).
This species has been confused with Periophthalmus koelreuteri (Pallas, 1770), itself a synonym of Periophthalmus barbarus (Linnaeus, 1766) according to Murdy (1989). P. barbarus does not occur in the Persian Gulf region. P. waltoni has been placed in the synonymy of P. koelreuteri in works on Persian Gulf fishes, e.g. by Relyea (1981), but the revision by Murdy (1989) is followed here. Another mudskipper species listed in Coad (1991b), Periophthalmus weberi Eggert, 1935, should be deleted from the list as Murdy (1989) does not report it from the Persian Gulf.
Two syntypes of Periophthalmus waltoni are in the Rijksmuseum van Natuurlijke Historie, Leiden (RMNH 17004) and 4 syntypes are in the Zoological Survey of India, Calcutta (possibly lost) (Murdy, 1989). However, this is more fish than listed in Koumans (1955).
Key characters
The pelvic fins are totally united into a sucker with a well-developed basal membrane or frenum (absent in P. barbarus). The anal fin base and second dorsal fin base are 27% or less of standard length and there are no canine teeth internal to the lower jaw symphysis in contrast to Boleophthalmus, the other mudskipper of southern Iran.
Morphology
First dorsal fin spines 10-14, second dorsal fin with 1 spine and 12-14 soft rays. Anal fin with 10-13 soft rays. Pectoral fin rays 13-15. Scales in lateral series 90-121, predorsal rows 27-37. The upper jaw has 4-5 anterior teeth on each side much larger than lateral teeth. Upper jaw teeth 19-28, lower jaw teeth 17-23. Gill rakers 9-16, mean 11, on each arch, triangular in shape with 3-4 spines at the top of each raker (Barak et al., 1994). The first dorsal fin lacks elongate spines and is slightly rounded. The dorsal fins are not connected by a membrane. The gut shortens with growth in terms of body length indicative of a carnivorous habit (Barak et al., 1994).
Sexual dimorphism
Females have a lozenge-shaped first dorsal fin (i.e. anterior 6 spines equal in height followed by abruptly shortened ones) with a height less than body depth; in males this fin equals body depth.
Colour
The overall colour is a light grey-brown. The flank bears blotches which, below the soft dorsal fin, take on the form of 8 short bars and there is a scattering of small silvery or white dots. Ventrally the spots form roughly transverse silvery stripes. The head top is also blotched and the sides bear a few small white dots. There are 7 irregular black blotches along the back. The first dorsal fin is dusky brown to light grey and its margin is yellowish between spines 1 to 8 and dark just below this except for the posteriormost 3-4 spines. There is a series of light spots on each interray membrane on the basal half of the fin. The second dorsal fin is dusky brown, with a pale margin, followed by a black stripe, and then with a white spot at the middle of each membrane thus forming a horizontal dotted line along the mid-fin. The caudal fin is a dusky brown, darker basally. There are 3 black blotches on the caudal peduncle. The pectoral fins are a light dusky brown on the rays with the membranes on the upper half pale with dark, brownish streaks in mid-fin, and a whitish margin is present. The pelvic fins are blackish with a reddish-brown or pale margin.
Size
Reaches 20.0 cm total length (Mhaisen and Al-Maliki, 1996).
Distribution
Found from the Persian Gulf to Pakistan. Reported from the Shatt al Arab, Iraq near Iranian Khuzestan by Khalaf (1961), Al-Nasiri and Hoda (1975b, 1976), Al-Hassan et al. (1989), and Hussain et al. (1989; 1997). Specimens believed to be this species have been reported from Iranian fresh waters but no specimens have been examined. Holly (1929b) records this species from the type locality of Barbus baschakirdi in the Bashakird Mountains (see Cyprinion watsoni). The Museum of Comparative Zoology, Harvard has a Periophthalmus species recorded from Manyuhi on the Shatt al Arab in irrigation ditches (MCZ 149602) but the identity of this specimen needs to be confirmed.
Abdoli (2000) maps this species from the Arvand River in the Tigris River basin, the lower Zohreh (= Hendijan), lower Helleh and Mand rivers in the Gulf basin, the Kul and Mehran rivers of the Hormuz basin, and the lower reaches of rivers of the Makran from the Jagin to the Bahu Kalat.
Zoogeography
This species has ready access to the sea and can colonise rivers along the Iranian coast.
Habitat
The habitat and behaviour of this species is similar to Boleophthalmus dussumieri (q.v.; Tytler and Vaughan (1983) review behaviour of this species in Kuwait, their Periophthalmus koelreuteri being this species according to Murdy (1989)). However it favours areas of mud which are drier on the upper and middle regions of the shore. The burrow is Y-shaped with two entrances and a depth of 30 cm in mud. Fish occupy several burrow systems within a 2-3 m home range (Clayton and Snowden, 2000). Young first appear on mud flats in April but stay in wet mud or shallow pools. They move onto drier areas by May after some growth (1.5-2.5 cm). There are up to 30 fish per 100 sq m on Kuwait mud flats. The activity range of body temperatures is 14-20°C in January and 26-34.5°C in June. Body temperatures do not exceed 35°C even when air temperatures exceed 40°C because of evaporative cooling. In tidal pools of a few centimetres depth in the Khawr az Zubayr, water temperatures ranged from 16 to 44ºC, pH from 7.6 to 8.7, dissolved oxygen from 5.2 to 8.8 mg/l and salinity from 10.2 to 20.2‰ (Al-Daham and Al-Noor, 2000). Holes from 50 to 80 cm depth, with two openings separated by not more than 50 cm, are constructed as refuge from extreme temperatures, high winds and predators. The mud is carried to the outside of the hole in the fish's mouth, and forms a dam-like structure around the shelter not exceeding 8 cm in height. The fish spends most of the ebb tide outside the shelter.
As well as being found on mudflats of the sea shore, this mudskipper is found in the Shatt al Arab and other rivers of the Persian Gulf.
Age and growth
Barak et al. (1994) studied this species in the Khawr az Zubayr, Iraq and found growth to be allometric with the regression coefficient significantly (p<0.05) greater than 3. (W = 1.74x10-4 L2.35 (r = 0.99)). Al-Daham and Al-Noor (2000) found the smallest mature males were 7.3 cm and females 7.4 cm in the Khawr az Zubayr with maturity at age one year. Sarafraz et al. (2011) found fish from Bandar-Pol and Qeshm Island had von Bertalanffy parameters L∞= 141.51 mm, k = 0.46 year-1 and t0 = 0.79 year and L∞= 166.79 mm, k = 0.42 year-1 and t0 = 0.62 year, respectively. Overall, males grew larger than females (208 mm versus 147 mm) while females grew faster (k = 0.41 versus 0.19 year-1). The age when the theoretical length was 0 (t0) was 1.46 year for males and 0.88 for females. Maximum age was estimated to be 4+ years.
Food
Fiddler crabs are stalked as the main prey of this mudskipper in Kuwait (Tytler and Vaughan, 1983) but shrimps and other invertebrates are also taken. High wind speeds and surface temperatures below 15ºC inhibited hunting. Hunts were spatially well separated because of the long time period before crabs returned to the surface after a hunt in their vicinity (Clayton and Snowden, 2000). On the intertidal mudflats of Iraq at the head of the Persian Gulf, this mudskipper feeds predominately on fish when larger than 90 mm (frequency 78.9%, occurrence 80.0%) with shrimps (14.1%, 20.0%), insects (3.2%, 10.0%), copepods 2.1%, 5.0%) and Daphnia sp. (1.7%, 5.0%). Fish smaller than 80 mm had values much lower for fish (6.7%, 8.3%) and higher for copepods (75.1%, 83.3%) with Daphnia sp. at 13.3% and 16.6%, insects at 2.7% and 4.1% and shrimps at 2.2% and 4.1%. Barak et al. (1994) found this species to be cannibalistic and piscivorous in the Khawr az Zubayr based on gill raker gap, gut-length/body length ratios decreasing with size, and stomach contents. Copepods were the major food item of smaller mudskippers (<70 mm) while mudskippers larger than this took fish. Mhaisen and Al-Maliki (1996) report food items as percentage of occurrence from fish caught in the Khawr az Zubayr estuary of Iraq to be 67.2% crustaceans (shrimps, crabs, barnacles and others), 34.5% snails, 3.4% insects and 1.7% fishes. Some fish consume plant material in winter months only, presumably in the absence of animal food.
Reproduction
Spawning in the marine environment of the Khawr az Zubayr takes place in March, the release of larvae being timed to take advantage of phytoplankton and zooplankton blooms (Hussain and Ahmed, 1999). Al-Daham and Al-Noor (2000) give the spawning period as March to April in the Khawr az Zubayr with a fecundity of 11,560-35,005 eggs for fish 7.4-12.9 cm long. A water temperature over 18ºC was the primary factor for spawning stimulation.
Parasites and predators
Mhaisen and Al-Maliki (1996) give information for Iraqi fish. E. Kahrom in the Crocodile Specialist Group Newsletter reports that muggers (marsh crocodile, Crocodylus palustris) eats a Periophthalmus in the Sarbaz River of Iranian Baluchestan (www.flmnh.ufl.edu/natsci/herpetology/newsletter/news173b.htm, downloaded 15 February 2002). However this may well be Boleophthalmus dussumieri.
Economic importance
None.
Conservation
No study of the numbers of this species in Iranian waters has been carried out so its status cannot be assessed.
Further work
The biology and population numbers of this species in Iran needs study.
Sources
The colour description is based on Kuronuma and Abe (1986).
Iranian material: None seen.
Comparative material: CMNFI 1985-0183, 2, 35.1-40.7 mm standard length, Iraq, Shatt al Arab (no other locality data); BM(NH) 1954.11.10:1-2, 2, 112.6-119.0 mm standard length, Iraq, Fao (29º58'N, 48º29'E); BM(NH) 1976.12.8:1-2, 76.1-84.1 mm standard length, Iraq, Basrah (30º30'N, 47"47'E); BM(NH) 1981.3.19:7-10, 4, 91.3-104.3 mm standard length, Kuwait mudflats (no other locality data).
This genus contains 13 species in in the Black and Caspian Sea basins (Freyhof,
2011) with 4 species in Iranian waters. Species were formerly included in
Neogobius but a molecular study by Neilsen and Stepien (2009a) separates
them.
Ponticola cyrius
Common names
gav mahi rud-e Kura (= Kura River goby) or gavmahi-ye Rud-e Kurosh.
[Kura goby, Kura River goby].
Systematics
Studies by Vasil'eva (1995a) and Vasil'yeva and Vasil'yev (1995) indicate that gobies from the upper and middle Kura River described as Gobius cyrius Kessler, 1874 and Gobius Weidemanni
Kessler, 1874, and identified by various authors as Gobius platyrostris, Gobius constructor, Gobius platyrostris cyrius, Gobius cephalarges constructor, Neogobius
cephalarges constructor and Neogobius platyrostris constructor should all be referred to
Neogobius (now Ponticola) cyrius.
Gobius cyrius was described from the Kura River near Borzhom, Georgia and 3 syntypes are in the Zoological Institute, St. Petersburg under ZISP 2235 (Eschmeyer et al., 1996). One of
these, a male 103 mm total length and 84 mm standard length, is designated as the lectotype by Vasil'yeva and Vasil'ev (1995). Two syntypes of Gobius Weidemanni described from Transcaucasia are
under ZISP 2224.
Key characters
The pelvic fin anterior membrane has large, angular lateral lobes, at least one-fifth width of rear edge and pelvic fin less than nine-tenths distance to anal fin; the posterior nostril is near the
edge of the orbit; lateral series scales usually 54-76; upper lip moderately swollen, width about 0.75 to more than length lateral preorbit; and interorbital distance 0.4-0.8 eye diameter.
Morphology
The head width is much greater than its depth. The interorbital distance is not more than half the eye diameter. The top of the head and the occiput are scaled with cycloid scales. The upper lip is
expanded towards the mouth corners, sometimes narrowing at the very end. The lower jaw protrudes and the angle of the jaws reaches back below the anterior half of the eye. The snout is longer than the
eye diameter (1.3-14 times). The upper lip is swollen at the angle. The second dorsal fin is of equal height along its length or rising slightly higher at its middle or end. The pelvic fin has a
well-developed membrane with pointed lobes and does not extend to the anal opening. The caudal peduncle is compressed, depth about 0.30-0.35 length. The caudal fin is usually rounded but may be pointed,
truncated or scalloped. The predorsal area is marbled and the first dorsal fin has a distal dark band.
First dorsal fin with 6-8, usually 6, spines, second dorsal fin with 1 spine and 15-20, usually 18-19, soft rays, and anal fin with 10-16, usually 14, soft rays (but see below), pectoral fin rays 17-19,
and lateral line scales 53-78. Vertebrae 33-34. The chromosome number is 2n=36-42 (Klinkhardt et al., 1995).
Meristics in Iranian material: first dorsal fin spines 6(38), second dorsal fin with 15(7), 16(28) or 17(3) soft rays, anal fin with 10(1), 11(8), 12(26) or 13(3) soft rays, and pectoral fin rays 17(3),
18(28) or 19(7). Lateral line scales 54(2), 55(5), 56(7), 57(11), 58(6), or 59(4).
Diploid chromosome number 36-42, chromosome arms 46, polymorphism seen in a pair of subteleocentric chromosomes, the number of which varies from 0 to 2.
Sexual dimorphism
Males may not become black during the breeding season (Vasil'yeva and Vasil'yev, 1995). There are various differences in morphometric characters, summarised in Vasil'eva and Vasil'ev in Miller (2003).
Colour
The body is marked by a reticulate or marbled pattern in dark fawn, by scattered light spots and by 8-10 elongate dark spots along the lateral line. The back anterior to the dorsal fin is marbled. The
belly is paler than the flank. The first dorsal fin is dark, particularly in the upper part where the darkening may form a band, with a transparent margin and a conspicuous black spot anteriorly
below the tip of the fin. The first dorsal, caudal and anal fins have light margins but the fins are mostly dark. The light edge is obvious in the anterior parts of the first dorsal and anal fins. Only the
second dorsal fin ray tips are light. The pectoral and pelvic fins are both uniformly dark.
Size
Attains 101.3 mm standard length, 13.0 cm total length.
Distribution
This species is found in the Caspian Sea basin including the Anzali Mordab and tributary rivers,
the Massuleh River at Siahbar, and the Pasikhan River just above confluence with
Pirbazar River and a tributary near Islamabad (Ahnelt and Holčík, 1996;
Abdoli and Naderi, 2009) and the upper and Middle Kura River and upper Aras River of the former U.S.S.R. (Berg, 1948-1949; Vasil'eva and Vasil'ev in Miller, 2003).
Zoogeography
This species is distributed in rivers of the Caspian Sea basin in Azerbaijan and Iran while Neogobius constructor, with which it has been confused, is found in the Black Sea basin. The absence
of P. cyrius from the lower Kura River could have been prevented by the prior penetration of
P. gorlap according to Vasil'eva and Vasilev (1995).
Vasil'eva and Vasilev (1995; 1996a) consider that Sarmatian marine ancestors of modern
P. cyrius and N. constructor most probably moved into fresh water during the Miocene, about 5 MYA.
Mountain building in the Caucasus isolated the Kura and other rivers which once flowed to the Black Sea and their course became oriented to the Caspian Sea. Isolation of the gobies resulted in speciation.
Ahnelt and Holčík (1996) present an alternative view. They place the origin of this species in the Quaternary when fresh water from melting glaciers connected the Black and Caspian seas via the
Manych Channel allowing freshwater fishes to disperse. This occurred four times during the Pleistocene, the last time about 14-12,000 years BP. A goby species was able to colonise both fresh and brackish
water and particular stocks began to favour certain habitats. Those stocks which favoured headwaters evolved into
P. cyrius while those which favoured lower regions and coastal areas evolved into
P. gorlap. Ahnelt and Holčík (1996) point out that there is an analogous situation in the Black Sea with N. constructor and the species N. rhodioni
Vasil'yeva and Vasil'yev, 1995. Additionally all west Caucasian and Caspian riverine gobies are very similar morphologically as evidenced by the difficulty in constructing reliable keys, suggestive of
recent differentiation about 11,000-9,000 years BP after the isolation of the Black and Caspian seas.
Habitat
This small goby is found only in freshwater in mountain and foothill zones in contrast to
P. gorlap. It is found in shallow (3 m or less) streams with a high gradient and current speed (1.6-4.9
m/sec on the bottom) in Iran (Ahnelt and Holčík, 1996). However, Vasil'yeva and Vasil'yev (1995) record it as absent in areas of strong current in the Kura River, preferring slow current and
muddy bottoms. This type of habitat is limited in Iranian rivers which are much shorter and smaller than those of the Caucasus where an extensive lowland course is found. In Iran, the stream bed is
composed of stones and boulders with some pebbles in the upper reaches and pebbles and gravel in the lower reaches. Small body size is an advantage for hiding under rocks to avoid predators and to escape
the rapid current.
Age and growth
Life span is up to 3 years with maturity attained in the second or third year (Vasil'eva and Vasil'ev in Miller, 2003).
Food
Vasil'eva and Vasil'ev in Miller (2003) report feeding on insect larvae and parts of plants.
Reproduction
This species may contain eggs in three size groups, the largest being 2.81 mm in diameter, indicative of batch spawning. Total fecundity reached 1060 eggs. Spawning occurs from May to August at 11-20ºC,
in some localities reported only for April-May (Vasil'eva and Vasil'ev in Miller, 2003).
Parasites and predators
Unknown.
Economic importance
None reported although in Azerbaijan some are caught by anglers.
Conservation
Further field work is necessary to assess numbers and status of this species.
Further work
The biology, distribution and conservation status of this species, recently recorded from Iran, remain to be examined in detail.
Sources
Counts and data are based on Ahnelt and Holčík (1996).
Ponticola goebelii
Common names
gavmahi-ye ratan, gel-ye mahi ratan. ?name cahnge [Kaspiiskii bychok-rotan or Caspian ratan or rotan goby in Russian].
Systematics
The Caspian Sea basin subspecies was usually referred to Neogobius ratan goebelii (Kessler, 1874). Gobius Goebelii
Kessler, 1874 was described from Baku, Azerbaijan and Gobius ratan was originally described from Odessa on the Black Sea.
Gobius Bogdanowi Kessler, 1874 described from the Caspian Sea at Makhachkala (Petrovsk) is a synonym (the male of N. ratan goebeli according to
Iljin (1956) although Pinchuk (1991) and
Ragimov (1998a) consider it to be a subspecies of N. ratan). Pinchuk (1976) notes that fish
from the Azerbaijan and Iranian coast differ from those from Dagestan and Turkmenistan, the first having fewer scales, but the number of specimens were insufficient to define taxa.
Gobius Goebelii has a syntype in the Naturhistorisches Museum Wien under NMW 33910 and others possibly in the Zoological Institute, St.
Petersburg (ZISP 2229-30) (Eschmeyer et al., 1996; Pinchuk et al., 2003). Two probable syntypes of Gobius ratan are in the Muséum national d'Histoire naturelle, Paris under MNHN A.1125 and a syntype is in the Zoologisches Museum Berlin (Museum für Naturkunde,
Universität Humboldt, Berlin) under ZMB 2098 (Bauchot et al., 1991).
Two syntypes of Gobius Bogdanowi are possibly in the Zoological Institute, St. Petersburg (ZISP 10902) (Eschmeyer et al., 1996; Pinchuk et al., 2003).
Key characters
Pelvic fin anterior membrane with angular lateral lobes, not more than one-fifth width of rear edge; pelvic fin almost reaches the anal fin (0.9 distance) or extends beyond the anal fin origin;
posterior nostril near edge of orbit; anterior nape with cycloid scales; rear of first dorsal fin without a dark spot; lateral series scales usually 49-54; and upper lip width 0.4-0.67 lateral preorbital
width (lip to orbit).
Morphology
Pelvic fin anterior membrane with angular lateral lobes, rather than rounded; lobes small, not more than 0.2 width of rear edge; posterior nostril near edge of orbit; angle of jaw below pupil of eye;
upper lip slightly swollen posteriorly, 0.5-0.67 lateral preorbital area (between lip and eye); first dorsal fin with sub-horizontal upper profile; rear of first dorsal fin without a dark spot; head depth
at eyes slightly less to somewhat greater than width between upper origins of opercles; snout length about 0.9-1.2 orbit; lateral series scales usually 52-61; interorbital width 0.4-0.6 eye diameter; nape
scales cycloid; pelvic fin almost reaches the anal fin (0.9 distance) or extends beyond the anal fin origin; the caudal peduncle depth almost equals its length, the posterior nostril is close to the eye
margin, and colour dark brown with few spots (spawning males black). The two subspecies listed above are distinguished partly by scale counts, 63-66 in bogdanowi and 63 or less in goebeli.
First dorsal fin with 5-7, usually 6, spines, second dorsal fin with 1 spine and 15-19 soft rays, anal fin with 1 spine and 11-15 soft rays, and pectoral rays 17-21. Range in lateral series scales
49-70. Vertebrae 32-34.
Sexual dimorphism
Male colouration is distinctive and they have a higher dorsal fin.
Colour
Overall colour is dark brown with a few, discrete, rounded pale spots and, along the mid-flank, a series of small horizontal bars. Pale saddles cross the back. The first dorsal fin has a yellow or
intense orange margin above a black bar (white in preserved fish). This fin also has a dark blotch, often with a bluish tint, on the first and second and second and third interradial membranes, and
sometime son the third and fourth interradial membrane. The second dorsal fin is greyish with 3-4 rows of brown spots proximally, and it may have a pale margin. The pectoral and caudal fins have rows of
rusty-brown spots proximally. The pelvic fin disc and the anal fin are grey with a broad pale margin. The belly is grey. Breeding males become black with a bluish tint.
Size
Reaches 23 cm.
Distribution
Found in the Black and Caspian seas including Iranian waters of the latter.
Zoogeography
See genus account above.
Habitat
This species lives in inshore waters over rocks, gravel and stones, although a spawning male was caught over silt near the Kura River, and a female was caught at 11 m at Krasnovodsk (Pinchuk
et al. in Miller, 2003). It rarely enters fresh waters, at least in the Caspian.
Age and growth
Sexual maturity is attained at 2 years.
Food
The main food items are crustaceans, with smaller quantities of worms, molluscs and fish.
Reproduction
Spawning takes place from the end of March to the end of May, with a single repeat spawning. Eggs are deposited on and between stones. Females contain 22-360 ripened eggs (Pinchuk et al. in
Miller, 2003).
Parasites and predators
None reported from Iran.
Economic importance
None as it is disliked by commercial and sport fishermen and is not common in the Caspian Sea (Pinchuk et al. in Miller, 2003).
Conservation
No data for Iran. Endangered in Turkey (Fricke et al., 2007).
Further work
The biology of this species in Iranian waters requires study.
Sources
Iranian material:
Ponticola gorlap
(Kessler, 1874)

(Kessler, 1874)

(Iljin, 1949)


Common names
mahi kafzi (= bottom-dwelling or benthic fish), gel-ye mahi sar bozorg (= bighead goby), gavmahi-ye sarbozorg, gav mahi sar gondeh, sebele, gorzak.
[Names once used for Neogobius kessleri gorlap may be applied to this species: iribogaz xul for Azerbaijan; bychok-golovach or bighead goby in Russian; gorlap goby, Caspian bighead goby]
Systematics
The Caspian Sea basin was thought to contain the subspecies Neogobius
(now Ponticola) kessleri gorlap (Iljin, 1949). Iljin (1956) and Vasil'yeva (1991) considered
P. k. gorlap to
be a distinct species while Berg (1948-1949) and Pinchuk (1977) on external morphological grounds considered it to be a subspecies. Karyotype data (Vasil'ev and Grigoryan, 1993) shows
P. k. kessleri
from the Black Sea basin to have 29 chromosomes in males and 30 in females and the number of biarmed chromosomes in 2n=29 individuals is 17 and in 2n=30 individuals it is 16. Nominal
P. k. gorlap
from the Caspian Sea have 2n=46 in both sexes (Vasil'yev and Vasil'yeva, 1992). Vasil'ev and Grigoryan (1993), Vasil'yev and Vasil'yeva (1992), Vasil'yeva et al. (1994) and Vasil'yeva and Vasil'yev
(1994) referred the Caspian Sea basin populations to Neogobius gorlap, but indicated that this taxon required redescription and this was carried out in 1996. Vasil'eva and Vasil'ev (1996) consider
that Gobius gorlap Iljin is unpublished and nonvalid, i.e. a nomen nudum (Iljin's 1941 manuscript is lost) and give the taxon the name Neogobius iljini with a type locality in the Mangyshlak
region of the Caspian Sea (holotype in the Zoological Museum of Moscow State University under ZMMGU P-19726 with paratypes ZMMGU P-46453 (3 specimens); note that the Zoological Museum of Moscow University
(ZMMU; their acronym) has P-19726 and P-4643 (sic)(Pavlinov and Borissenko, 2001)). Kottelat (1997) points out that Berg (1948-1949; volume 3 including gobies dated 1949) had access to and used some
of Iljin's data, thus making the name gorlap available. Indeed, any of the subsequent papers by Vasil'eva and Vasil'ev and others using the name with descriptive details would have made the name
available. Neogobius iljini is therefore a junior synonym of Ponticola gorlap
(Iljin in Berg, 1949). There is no type series for P. gorlap as it is based on
Iljin's data as outlined
above.
Key characters
Pelvic fin anterior membrane with angular lateral lobes, lobes about one-sixth to almost one-half width of anterior edge of membrane; pelvic fin less than nine-tenths distance to anal fin; posterior
nostril near edge of orbit; anterior nape with ctenoid or cycloid scales; rear of first dorsal fin without a dark spot; lateral series scales usually 54-76; lateral lobes of pelvic fin anterior membrane
large, at least one-fifth width of rear edge; upper lip not markedly swollen, width at least 0.6 lateral preorbital width (lip to orbit), if less than 0.75, then nape scales ctenoid; interorbital distance
0.8-0.9 eye diameter; and caudal peduncle depth 0.67-0.75 length.
Morphology
This species is separated from other Caspian gobies in Iran by having a completely scaled nape, scales usually cycloid on the sinciput (anteriorly on the head just behind the eyes) (70-100%
cycloid; 0-30% uniseriate ctenoid in Kura River fish) and cycloid or ctenoid posteriorly over the operculum (60-100% ctenoid in Kura River fish, 0-50% ctenoid in others); scales on the middle of the upper
third of the operculum usually cycloid (70-100%); head depth at the eyes is slightly less or about equal to head width between the upper origins of the opercles; a large mouth with the jaw angle below the
anterior part of the eye; the snout is much longer than the eye, 1.5-1.6 times; upper lip not expanded, or only slightly, less than lateral preorbital area (between lip and eye); interorbital distance less
than 0.85 eye diameter; first dorsal fin not higher than second dorsal fin; moderately large, angular or pointed rather than rounded lateral lobes to the anterior pelvic membrane (rather small and obtuse
lobes according to Ahnelt and Holčík (1996)), the membrane is 0.2-0.3 the width of the rear edge as opposed to not more than one-sixth; pelvic disc length is about 0.8 length of pelvic origin to anus
length; scales in lateral series 53-79; caudal peduncle depth is 0.6-0.75 its length; the posterior nostril is close to the eye margin; and overall colour is yellowish and marbled.
The head in fish from the Kura River and possibly Iran have a rather subcylindrical shape (43.8-60.0%), while marine and north and middle Caspian Sea basin fish have a depressed and wide head, the width
being markedly greater than the depth. The caudal fin is rounded, oval, or rarely elongated.
First dorsal fin spines 5-7, usually 6, second dorsal fin with 1 spine followed by 15-20, usually 18, soft rays, anal fin with 1 spine and 11-16, usually 13-15 (but see below), soft rays, pectoral fin
rays 17-22, and pelvic fin rays 8-9. Marine specimens and those from the lower Volga River and Yashan Lake in the Uzboi River system of Turkmenistan have less than 15 anal fin branched rays while
freshwater populations (in the Kura River at least, perhaps Iran) have 30-60% of all fish with 15 rays. Scale counts in the southern Caspian Sea are less (53-74) on average than for other populations
(61-79) (Vasil'eva and Vasil'ev, 1996). Gill rakers 5-8. Vertebrae 33-35. The chromosome number for
P. gorlap is distinctive with 2n=46 with 46 chromosomal arms (Vasil'ev and Grigoryan, 1993;
Esmaily and Kalbassi, 2008).
Meristic values for Iranian specimens are: first dorsal fin spines 6(16), second dorsal fin soft rays 16(4), 17(11) or 18(1), anal fin soft rays 12(13) or 13(3), pectoral fin rays 18(8) or 19(8) and
lateral line scales 63(5), 64(2), 65(2), 66(2), 67(2) or 68(2).
Sexual dimorphism
Abdurakhmanov (1962) reports on fish from Azerbaijan that head width, predorsal distance, second dorsal fin and anal fin bases and length of third branched anal fin ray are all greater in males while
pelvic fin length and eye diameter are greater in females. In sea populations, females are larger than males.
Colour
A strongly yellow to reddish-yellow coloration distinguishes this species (Berg, 1948-1949; Vasil'eva and Vasil'ev, 1996) although Gavlena (1977) describes Volgograd Reservoir specimens as brown and
others agree fish may be dark brown to dark grey or greenish. There are 4-8 elongated dark brown spots along the mid-flank in freshwater specimens, the last forming a horizontal "T" with the
cross-bar on the caudal fin base. The flanks are usually marbled dark brown with light brown and yellowish strips, sometimes in a cellular design. There are light spots, darkly rimmed, along the sides of
the head and near the pectoral fin bases, the latter also being likened to a network, reticulate pattern or cellular design. Cheeks with reticulate or roundish-cellular design. Short, longitudinal stripes
often present below the eyes. The belly is light grey or light yellowish.
Dorsal, caudal and pectoral fins yellowish to light brownish. Second dorsal fin branched ray tips white. The fins typically have rows of small dark, orange-grey spots, although the pelvic fin has only
a little dark grey or yellowish pigment with some brownish strips or many minute spots. The first dorsal fin lacks a large spot but has 3-4 series of spots; the second dorsal fin has 2-6, usually 3-4,
series of spots; the anal fin is yellow to grey with occasional spots; the pectoral fin has 8-10 series of spots; and the caudal fin has 7-8 series of spots. The ends of the second dorsal fin branched rays
are white. The anal fin has a light border as does the pelvic disc, although the latter sometimes has a black centre. The caudal fin has many minute spots. The genital papilla is dark grey. Young fish may
appear spotted but have a more reticulate pattern on the flank than N.
goebelii and are generally darker.
The dorsal fins in breeding males have a wide yellowish or white margin and the second dorsal fin rays have free tips. The margin is rusty in a female. The head can be a dark bluish-grey and the flanks
are dark with some paler markings. Fin membranes become dark.
Size
Reaches 22.5 cm. Freshwater populations have fish much smaller than those in the sea (Vasil'eva and Vasil'ev in Miller, 2003).
Distribution
Found in the inshore Caspian Sea and tributary rivers. It is reported from between Kultuk and Astara in Azerbaijan (Ragimov, 1965) and from the Uzboi Lakes in Turkmenistan. In Iran, it is reported
from a wide range of rivers from Astara to the Gorgan and probably the Atrak,
including the Siazan, Safid, Duyusan, Shimrud, Khoshkerud, Palarud, Naviq,
Shasdehrud, Shafarud, Kargan, Gharasu, Tajan, Babol, Haraz, Pol-e Rud and Safid,
the Aras River, Bahambar, Siah Darvishan, Pasikhan, Massouleh and Pir Bazar rivers,
Anzali Mordab and Shore, Gorgan Bay, the southeast Caspian Sea, southwest Caspian Sea and south-central Caspian
Sea (Holčík and Oláh, 1992; Ahnelt and Holčík, 1996; Abbasi et al., 1999; Kiabi et al., 1999;
Abdoli and Naderi, 2009; K. Abbasi - see photographs above).
Abdoli and Naderi (2009)
consider it to be absent from the Atrak, Sardab, Tonekabon and Aras rivers. Zoogeography
This species is thought to have evolved from populations which settled in the lower reaches of rivers and in coastal areas while those populations which penetrated headwaters evolved into
P.
cyrius (q.v.) (Ahnelt and Holčík, 1996). See also genus account above.
Habitat
This species is found in fresh and brackish waters in rivers and lagoons and in inshore waters of the Caspian Sea. In the sea, it is found down to 10 m, rarely to 20 m, on rocky and dense sand bottoms.
It moves inshore in the sea for spawning and retreats to deeper water (6-12 m) to overwinter (Vasil'eva and Vasil'ev in Miller, 2003). It may be found among reeds. It can live at river sites with
a current velocity at 4.5 m/sec on the bottom (Holčík and Oláh, 1992). This species is allopatrically distributed with
Ponticola cyrius (q.v.). Ahnelt and Holčík (1996) rate
this species as a eurytope, inhabiting both fresh and brackish waters, slow and rapid currents, and clay and pebble and gravel bottoms.
Kirilenko and Shemonaev (2010) found fish in the Kuibyshev Reservoir in Russia
to prefer rocky, sandy-slimy grounds rich in higher aquatic vegetation and are
sometimes found among cane roots. Age and growth
Brackish water populations grow larger than freshwater ones (Ahnelt and Holčík, 1996). Males attain 3 years, females 2 years, and males grow faster than females. Maturity is reached at
1+ for most females and 2+ years for males (Vasil'eva and Vasil'ev in Miller, 2003).
Kirilenko and Shemonaev (2010) found a female in the Kuibyshev Reservoir that
was 4 years old while the oldest fish in the 3+ class were all males, fish
matured at 1+ years for both sexes. Abdoli et al. (2006) examined fish
from the Gomishan and Miankaleh wetlands and found a length (L)-weight (W)
relationship of W = 0.1L2.39 for males, 0.0595L2.588 for
females and 0.1L2.4 for sexes combined. Food
The principal food in the sea is fishes, mostly gobies and Atherina, but also includes crustaceans. Benthic food means this species competes with commercial fishes. In fresh water, fishes eaten
include Nemacheilidae and the cyprinid Alburnoides bipunctatus (=
eichwaldii), as well as crustaceans such as gammarids (e.g. Dikerogammarus haemobaphes), gobies such as its own species and N.
melanostomus, insects and clams (Gavlena, 1977). Kirilenko and Shemonaev
(2010) found food in the Kuibyshev Reservoir to be mainly crustaceans and
chironomids. Reproduction
Spawning takes place in June in the Volgograd Reservoir and each female may contain up to 576 eggs (Gavlena, 1977). In the Caspian Sea, fish have as many as 3506 large eggs and 3399 smaller ones.
Spawning takes place on rock and shingle bottoms with eggs laid on various objects. The males of this species are nest guarders. Spawning in the southern Caspian is in April and May with two batches of
eggs deposited. In the lower Volga River, spawning is from the end of April to July (Vavsil'eva and Vasil'ev in Miller, 2003).
Parasites and predators
The Caspian seal, Pusa caspica, is a predator on this species (Krylov, 1984). Pazooki and Aghlmandi (1998) found the nematode Dichelyne minutus infecting the intestine of 100% of
fish examined from the south Caspian Sea in Iran. and Sattari et al.
(2002; 2004) and Sattari (2004) record the presence of the nematode, Eustrongylides excisus, in this species. This parasite can damage muscles in commercial
species and render them unsuitable for sale. Dichelyne minutus was also
recorded. Daghigh Roohi and Sattari (2004)
record Eustrongylides excisus, Dichelyne minutus and Corynosoma
strumosum from this species in the southwestern Caspian Sea of Iran. Sattari et al. (2005) surveyed this species in the inshore area of the Caspian Sea, recording Eustrongyloides excisus and Dichelyne
minutus.
Economic importance
None in Iran although it has formed a small part of the goby catches in the former Soviet Union. It is also caught by anglers (Vasil'eva and Vasil'ev in Miller, 2003).
Conservation
Kiabi et al. (1999) consider this species to be of least concern in the south Caspian Sea basin according to IUCN criteria. Criteria include abundant in numbers, widespread range (75% of water
bodies), absent in other water bodies in Iran, and present outside the Caspian Sea basin.
Further work
The biology of this specie sin Iran needs work.
Sources
Meristics and colour description based in part on Gavlena (1977) describing fish from the Volgograd Reservoir, identified as N. kessleri. Counts and data for Iranian fish are based on Ahnelt and
Holčík (1996).
Iranian material:
Ponticola syrman
Common names
None (other than general names listed under genus above).
[shirman in the Ukraine, syrman goby, Kaspiiskii shirman or Caspian syrman goby]
Systematics
Gobius syrman is described from Odessa, Dniester estuary, in the former U.S.S.R., now Ukraine). Gobius Trautvetteri Kessler, 1859 described from the lower Bug and Dniester rivers in the
former U.S.S.R. and Gobius eurystomus Kessler, 1877 described from the southern Caspian Sea are synonyms. Berg (1948-1949) has eurystomus as a valid subspecies of Neogobius syrman but this
is not accepted now (Reshetnikov et al., 1997). Presumed syntypes of G. syrman are in the Natural History Museum, London (BM(NH) 1872.5.30:35, 105.0 mm standard length), Muséum national
d'Histoire naturelle, Paris (MNHN A. 1126) and in the Naturhistorisches Museum Wien (MNMW 30099. Syntypes of G. eurystomus are under BM(NH) 1897.7.5.8-9 (2, 73.6-88.6 mm standard length),
in the Zoological Institute, St. Petersburg (ZISP 10904-05), and perhaps NMW 29176. (Pinchuk et al. in Miller, 2003).
Key characters
Three transverse suborbital neuromasts rows occur ventral to the longitudinal suborbital row b while all other Neogobius have typically two rows.
Additionally, pelvic fin anterior membrane with rounded and shallow lateral lobes, lobes not more than one-sixth width of anterior edge of membrane, or lacking entirely; pelvic fin disc two-thirds
distance to anal fin origin; nape with ctenoid scales; first dorsal fin without large dark spot; lateral series scales usually 55-70; and angle of jaw below pupil of eye.
Morphology
First dorsal fin with 5-7, usually 6, spines, second dorsal fin with 1 spine and 15-19, usually16-18 soft rays, anal fin with 1 spine and 10-15, usually 12-14, soft rays, and pectoral rays 17-21.
Range in lateral series scales 56-79. Vertebrae 34-35. Pelvic fin anterior membrane with very shallow, rounded lateral lobes. Posterior nostril near edge of orbit. Angle of jaw below anterior part of
pupil, upper lip with uniform width, about half lateral preorbital area (between lip and eye). First dorsal fin relatively high, with rounded upper profile, without an upper anterior dark blotch. Second
dorsal fin highest in middle. Head depth at eyes 1.1-1.2 width between upper origins of opercles. Interorbital width less than or equal to eye diameter. Snout length 1.3-1.4 orbit. Lateral series scales
usually 59-67. Nape and predorsal area scaled completely. Breast, base of pectoral fin and upper part of opercle scaled, cycloid scales on ventral part of abdomen, opercle and anterior part of nape.
Pelvic fin about two-thirds distance to anal fin origin. Caudal peduncle depth about half length. Posterior nostril close to the eye margin. The head canals and the free neuromasts on the body are
typically arranged for a neogobiine. Three transverse suborbital neuromasts rows occur ventral to the longitudinal suborbital row b.
Sexual dimorphism
Males may have a more intensely dark distal band and body spots during spawning. Male median fin rays are slightly elongated during spawning. Females are usually smaller than males.
Colour
Overall colour pale grey, brownish-grey or more rarely yellowish-brown (perhaps fish parasitised with the strigeid nematode Neascus), with pale saddles across the back extending onto the flank
and sometimes meeting dark blotches along the mid-flank. The body has large grey-brown markings arranged in a chequer-like pattern. The head has brown markings laterally and a dark band antero-ventrally
from the eye to the upper lip. The first dorsal fin has a pale yellowish margin set off by a dark stripe above two brown stripes proximally. The second dorsal fin bands are less distinct and the anal fin
has an indistinct, bluish band and a white margin. The pectoral fins are yellowish and the pelvic disc is pale. Belly pale. Males do not become dark during spawning (Pinchuk et al. in Miller, 2003).
Size
Attains 29.2 cm.
Distribution
Black and Caspian Sea basins including the Imeni Kirova or Kyzylagach Bay near the northwestern Iranian border (Kuliev, 1989) and between Kultuk and Astara in Azerbaijan (Ragimov, 1965) and recently
recorded from Iran (Ahnelt et al., 2007). Reported from the Anzali Mordab and Gomishan Lagoon,
Gorgan Bay, and the southwestern, southeastern and south-central Caspian Sea (Jolodar and Abdoli, 2004;
Abdoli and Naderi, 2009). Introduced to the Aral Sea.
Zoogeography
See genus account above.
Habitat
This goby is found principally in the sea, with one report from fresh water in the Caspian Sea basin (Emba River, Kazakhstan). It is unusual in being found on mud and silt bottoms and can tolerate low
oxygen concentrations (to 20% saturation). It is rare offshore at 6-20 m in the Caspian Sea. It is found in the Caspian at 1-10 m, retreating in summer to slightly deeper water, reappearing as autumn
cooling occurs, and then spending the winter at 30-50 m. It approaches shores earlier than other gobies for spawning, as early as the latter half of February for males with females in the second half of
March (Pinchuk et al. in Miller, 2003).
Age and growth
Maximum age may be 6 years although most fish probably live only half this time. Maturity may be attained shortly before age 1 year and by 2 years for most fish.
Abdoli et al. (2006) examined fish from the Gomishan and Miankaleh
wetlands and found a length (L)-weight (W) relationship of W = 0.0033L3.41
for males, 0.0033L3.5 for females and 0.003L3.5 for sexes
combined. Food
Fish, crustaceans and Nereis worms are the main diet items in Caspian Sea fish examined. Fish included Knipowitschia and fry of Neogobius
pallasi and N. melanostomus as
well as clupeids and its own species. Smaller individuals take crustaceans and larger ones favour fish (Pinchuk et al. in Miller, 2003).
Abdoli et al. (2012) found fish in the Gomishan Wetland feeding on
chironomids, amphipods and fish, preferring fish in autumn and summer and
chironomids in spring and winter. Reproduction
The spawning season is from the beginning of April into May in the southern Caspian Sea. The eggs are found in two sizes, indicative of batch spawning. Up to 8474 ripe eggs are found in each female.
Eggs are 4.2 by 1.9 mm and are laid under and between stones, on any available object or even muddy bottoms.
Parasites and predators
This species is eaten by Sander lucioperca and fry are eaten by sturgeons.
Economic importance
This species has been fished in former Soviet waters of the Caspian Sea because of its large size but the proportion in catches varies from 1-50%. It is important as food for commercial species.
Conservation
Only recently recorded from Iran, its conservation status in unknown.
Further work
The distribution and abundance of this species in Iranian waters needs to be surveyed as it is an important food for commercial fishes.
Sources
Pinchuk et al. in Miller (2003) is a recent summary of data on this species.
Iranian material: CMNFI 1970-0544, 1, 141.7 mm standard length, Gilan, near Bandar-e Anzali, (37°28’N, 49°27’E);
CMNFI 1979-0689, 9, 87.1-124.0 mm standard length, Gilan, Safid River at Hasan Kiadeh (37°24’N, 49°58’E);
CMNFI 1970-0586, 1, 100.1 mm, Mazandaran, Gorgan Mordab at Ashuradeh-ye Kuchak (36°50’N, 53°56’E);
NMW 95074, 2, 100.1–130.2 mm, Mazandaran, south-east Caspian Sea near estuary of Gorgan River (36°59’N, 53°59’E);
CMNFI 1979-0788, 2, 135.3–146.2 mm standard length, Mazandaran, Gorgan River (37°00’N, 54°07’E);
NMW 95075, 1, 186.6 mm standard length, Mazandaran, south-east Caspian Sea, Mian Kaleh near Behshahr (36°53’N, 53°32’E).
Genus Proterorhinus This genus is characterised by having 6 first dorsal fin rays, a
pelvic disc without obvious lateral lobes and, uniquely in Ponto-Caspian gobies, a very elongate
anterior nostril which hangs over the lip. Miller in Miller (2004) give
additional characters, in particular the distribution of neuromasts on the head. It contains only a single
species found in the Black and Caspian seas and their tributary rivers
and in the northern Aegean Sea in Greek fresh waters. There may be two species
if P. semilunaris is recognised (see below). Simonovič (1999) considered this genus to be a
subgenus of Neogobius but Ahnelt and Duchkowitsch (2001) provisionally
consider Proterorhinus to be distinct based on their study of the lateral
line system and consider it distinct based on osteology (Ahnelt and Duchkowitsch,
2004). Miller in Miller (2004) recognises the genus as distinct as do Stepien
and Tumeo (2000) based on mtDNA. Simonovič et al. (1996)
consider this genus of gobies to be a young one as evidenced by the distribution of the species in the Caspian Sea which closed recently.
Reshetnikov et al. (1997) and Miller in Miller (2004) have the date for this genus as
1899 but Eschmeyer's "Catalog of Fishes" states that it was published in 1900 although dated 1899.
Proterorhinus nasalis
Common names
gav mahi, sag mahi, gel-ye mahi marmari, gavmahi-ye marmari, sebele.
[marmar xul in Azerbaijan; bychok-tsutsik or tubenose goby,
marmornyi bychok or marbled goby, both in Russian; mottled goby].
Systematics
The Caspian Sea tubenose goby has generally been referred to Proterorhinus marmoratus
(Pallas, 18l4). Gobius marmoratus was originally described from Sevastopol, Ukraine, on the Black Sea.
Types are lost (Miller in Miller, 2004).
Gobius nasalis De Filippi, 1863 described in Latin from the
"Mar Caspio presso Baku" and Gobius blennioides
Kessler, 1877 described from "Bakinskoi bukhte" (= Baku
Bay), were regarded as synonyms. The meristic characters reported by De Filippi
(1863) for nasalis are probably in error according to Berg
(1948-1949) but the only character falling outside the ranges compiled
below is for soft dorsal rays given as 21.
Stepien and Tumeo (2006) used mtDNA cytochrome b sequence data and
consider there are two species in this genus. P. marmoratus is a marine
species in the Black Sea and P. semilunaris (Heckel, 1837) is a
freshwater species in other Eurasian habitats. The type locality of the latter
is "Maritza [Marizza] R., near Plovdiv, e. Rumelia, Balkan region of Bulgaria".
Caspian Sea basin Proterorhinus were not investigated. Kottelat and Freyhof (2007), Freyhof and Naseka (2007) and
Sorokin et al. (2011) restrict marmoratus to brackish waters in
Sevastopol and consider nasalis to be the name for Iranian Caspian Sea
populations.
Neilson and Stepien (2009b) examined mtDNA and nuclear DNA and confirmed the
presence of a distinct taxon in the Caspian Sea basin. They advocated P. semipellucidus
as the name for the Caspian Sea basin taxon. Gobius semipellucidus
Kessler, 1877 was
described from a single specimen taken at the "ust'e rechki Kara-su,
vpadayushchei vb Astrabadskii zalivb" (= mouth of the Karasu
River falling into Astrabad Bay (Qareh Su, Gorgan Bay) and the
holotype is in the Zoological Institute, St. Petersburg (ZISP) (Eschmeyer
et al., 1996). P. semipellucidus is given as a valid species in Eschmeyer's "Catalog of
Fishes" but is presumably a synonym of P. nasalis. The relationship
between marine Caspian Sea (brackish) populations (nasalis) and true
freshwater populations (possibly semipellucidus) remains to be resolved.
Neilson and Stepien (2009b) examined only freshwater and brackish (0-6 p.p.t.)
specimens from the Caspian Sea basin. Sorokin et al. (2011) consider
nasalis to be the taxon widely distributed in the Caspian and Sea of Azov
basins with pellucidus a synonym, although further work is needed to
determine if pellucidus is the freshwater taxon. The syntypes of Gobius nasalis are in the Istituto e Museo
di Zoologia della R. Università di Torino under MZUT N.672 (7
specimens), the Natural History Museum, London under BM(NH)
1869.3.4:34 (1, 43.4 mm standard length), in the Museo Civico di Storia Naturale di Genova
under MSNG 12655 (2) and MSNG 36228 (3), in the Naturhistorisches
Museum Wien under NMW 33894 (1), NMW 33895 (1) and NMW 33896 (1), and
in the Museum für Naturkunde, Universität Humboldt, Berlin under ZMB
5015 (3) (Eschmeyer et al., 1996; Miller in Miller, 2004; 43.3-48.4 mm
standard length, measured in February 2006). The holotype of Gobius semipellucidus
is lost. Ahnelt and Mikschi (2008)
give details on the types of Gobius semilunaris. Key characters
The combination of the pelvic fins forming a disc, the head not
being flattened, and the anterior nostrils forming a tube overhanging
the upper lip are distinctive. The origin of the first dorsal fin is displaced
anteriorly in comparison to other Ponto-Caspian and Atlantic-Mediterranean
gobioids, and this reflects the differing postcranial osteology detailed in Ahnelt and Duchkowitsch (2004). Morphology
First dorsal fin spines 5-7, usually 6, second dorsal fin with 1
spine and 12-20, usually 16-17, soft rays, anal fin with 1-2 spines and 11-17,
usually 13-14, soft rays, and pectoral fin with 14-16 branched rays. Scales in lateral
series 34-53. Scales are a vertical oval with the anterior margin
rounded to slightly wavy, the upper and lower margins rounded and the
posterior margin with almost straight upper and lower elements coming
almost to a central point. This causes the exposed portion of the
scales to be delimited by almost straight edges and flank scales have
a diamond pattern. Circuli are numerous and fine. Radii are numerous
and radiate back from a focus almost at the posterior margin. The
posterior scale margin bears fine ctenii. Gill rakers 3-6, short and
concentrated around the angle of the gill arch. A membrane partly
closes off and narrows the gill opening at the gill raker level and
may partially encompass a raker making counts difficult. Vertebrae
30-33. The chromosome number is 2n=46, with 46 acrocentric chromosomes
and 46 chromosomal arms (Ráb, 1985; Grigoryan and Vasil'ev, 1993; Simonovič, 1999).
The predorsal area on the back and the nape are scaled. The cheek has
infraorbital neuromast organs in 7 transverse rows from the lower orbit border
without uniserial papillae between the rows. The preopercular canal
has three pores, the anterior oculoscapular canal has 7 pores
and the posterior oculoscapular canal has 2 pores. Ahnelt
and Duchkowitsch (2001) give a detailed description of the lateral line
system on the head. The gut is short and s-shaped. The genital papilla has a two-lobed tip.
Freyhof and Naseka (2007) note that Caspian tubenose gobies are most similar
to P. semilunaris (Heckel, 1837) of the Aegean and Black seas basins and
are distinguished by having a shorter posterior membrane to the first dorsal
fin, not reaching the origin of the second dorsal fin (versus reaching), a
longer anterior naris reaching to the middle or to the lower margin of the lower
lip when folded down (versus to the upper lip or to the upppermost margin of the
lower lip), and a smaller eye (diameter 16-21% head length versus 20-28%). Meristic values for Iranian specimens are:- first dorsal fin spines 5(1),
6(28) or 7(1); second dorsal fin soft rays 13(1), 14(6), 15(17) or
16(6); anal fin soft rays 12(13), 13(14), 14(2) or 15(1); total
pectoral fin rays 14(18) or 15(12); lateral series scales 36(1),
38(4), 39(7), 40(7), 41(4), 42(5), or 44(2); total gill rakers 3(18),
4(9) or 5(3), although accuracy is doubtful because of the membrane;
and total vertebrae 31(12) or 32(13) (courtesy of Nina Bogutskaya, January 2012).
Sexual dimorphism
Males are overall darker than females, e.g. one at 60.1 mm standard
length had pigment on both rays and membranes of fins, and all fin
margins frilly and pale or orange-yellow. Cheeks are swollen in spawning males
and fins are longer than in females.
Colour
Overall colour is brownish or yellowish-brown to olive or yellowish-green with about
5 oblique and irregular bars or blotches across the flank. Colour varies with
the background, being darker on mud bottoms and a bright reddish-brown on sand. The head
has brown-grey bands over the top and sides on a pale background. The
first dorsal fin may have a pinkish tinge and irregular grey blotches.
The second dorsal fin has several series of brown spots with the upper
2-3 greenish-yellow. The caudal fin is grey interrupted by up to 9
narrow bands of greenish-yellow. There is a triangular black spot at
the caudal fin base, flanked by two white spots. The pectoral fin is
similarly greyish with 5-7 narrow bands of pale green or yellow. The
pelvic fin has 3-4 yellow bands and the anal fin 6-7 yellowish and
oblique bands over a grey background. The iris has a narrow golden
ring. Spawning males have all fins dark, although the pectoral fin is
lighter than the rest and the caudal fin has an obvious white margin.
The genital papilla is grey. Breeding females have a colour similar to
that of both sexes in winter. Young (16-22 mm) have an unpigmented
median band, wider than the eye, from the head to the first dorsal fin
and even to the caudal fin. In front of the first dorsal fin is a dark
saddle. The peritoneum is light brown to silvery with varying amounts of melanophores.
Size
Attains 15.0 cm although Caspian specimens reputedly reach only 7.6 cm.
Distribution
Found in the Caspian Sea basin and the Uzboi Valley of Turkmenistan. It is found in a wide range of rivers on the Caspian coast of Iran
including the Qareh Su falling into Gorgan Bay, Kargan River estuary near
Hashtpar, lower Safid, Rud-e Sera and Babol, in the Anzali Mordab and
Kolesar River, the southeast Caspian Sea,
the southwest Caspian Sea and south-central Caspian Sea (Derzhavin, 1934; Holčík and Oláh,
1992; Abbasi et al., 1999; Kiabi et al., 1999;
Abdoli and Naderi, 2009).
Zoogeography
The genus Proterorhinus diverged from the related
genus Neogobius about 5.2MYA during the late Miocene/early
Pliocene according to mitochondrial DNA evidence (Dillon and Stepien, 2001).
Freshwater and marine/brackish populations apparently diverged 3.82-4.30 MYA and
freshwater lineages in the Black and Caspian seas diverged 0.92-1.03 MYA
(Neilson and Stepien, 2009b). Habitat
This species occurs in both salt and brackish waters and enters the
fresh water of rivers. There are some permanent freshwater populations
in Europe. The usual habitat is shallow sea bays, offshore banks or flowing water
of streams but it may also be found
in ponds and canals overgrown with vegetation. Where current is strong it hides
under boulders. Some fish are found below 3 m depths in the sea. It is often found under
stones or among weed, to which it can retreat rapidly if disturbed.
Most activity takes place at night.
Age and growth
Life span is variably reported as 2 or about 5 years with maturity attained at 1-3 years.
Young-of-the-year can mature by autumn at 5.5 cm in the Caspian Sea (Miller in Miller, 2004).
Food
Kuliev (1989) records insect remains, gammarids, daphnia,
chironomids, molluscs and fish eggs in the guts of this species in
Kirova Bay of Azerbaijan. For the Caspian Sea as a whole, amphipods
predominate at 45.7% followed by mysids at 21.8%, cumaceans at 16.4%
and decapods at 7.2%. Molluscs make up 5.6% (Kosarev and Yablonskaya,
1994). Iranian specimens contained crustacean and insect remains. Elsewhere
polychaete worms and small fish have been found in stomachs of this species
(Miller in Miller, 2004).
Reproduction
Both sexes produce sounds during the breeding season. Eggs are laid from shallow water (0.5 m) down to 20 m in
bowl-shaped nests, under rocks, in empty mollusc shells, and on roots
and stems of plants. Empty cans may be used. Two females may lay eggs in the
same nest. Spawning begins in mid-April in the southern Caspian and may continue as late as the
first half of August. Some fish may be mature in the middle of March
in Iranian waters. Fish presumed to be young-of-the-year at 9.8 mm
standard length have been collected on 26 April, by 10-11 June fish
are 11.2-11.6 mm (mean 11.4 mm), on 20 June the 5 smallest fish of a
sample were 12.5-14.1 mm, mean 13.6 mm while the 5 largest were
21.0-25.2 mm, mean 23.4 mm, and on 12 October the 5 smallest were
16.6-19.9 mm, mean 17.7 mm while the 5 largest were 26.4-33.3 mm, mean
29.6 mm. By 15 March, eggs in a 49.7 mm standard length fish are 1.3
mm in diameter and well-developed. This data suggests an extended
spawning season in Iran. However young are first found in mid-June
according to Ragimov (1987). Eggs are deposited in 2-3 batches and are
guarded. Females contain up to 1335 eggs of diameter 3.3-3.6 by 1.4-1.5 mm.
Elsewhere up to 2500 eggs may be laid. Egg clutches can be transported
on the hulls of ships (Ahnelt et al., 1998). Males are nest guarders.
Parasites and predators
Unknown for Iran.
Economic importance
Robins et al. (1991) list this species as important to North
Americans. Importance is based on its use in aquaria and because it
has been introduced outside its natural range (Jude, 2001).
Conservation
Lelek (1987) classifies this species as vulnerable in Europe on
account of water level changes. Kiabi et al. (1999) consider
this species to be data deficient in the south Caspian Sea basin
according to IUCN criteria. Criteria include few in numbers, medium
range (25-75% of water bodies), absent in other water bodies in Iran,
and present outside the Caspian Sea basin.
Further work
The biology of this species in Iran is not well known.
Sources
Type material: See above for Gobius nasalis (ZMB 5015).
Iranian material: CMNFI 1970-0530, 66, 41.6-64.5 mm standard length, Gilan, Nahang Roga River (37º28'N, 49º28'E);
CMNFI 1970-0542, 5, 48.0-59.6 mm standard length, Gilan, Old Safid River estuary (37º23'N, 50º11'E);
CMNFI 1970-0585, 2, 52.6-55.9 mm standard length, Gilan, Nahang Roga River (37º28'N, 49º28'E);
CMNFI 1970-0590, 5, 39.5-47.7 mm standard length, Mazandaran, Shesh Deh River near Babol Sar (no other locality data);
CMNFI 1979-0787, 3, 52.7-59.1 mm standard length, Gilan, Nahang Roga River (37º28'N, 49º28'E).

Anzali Shore, December 2009, courtesy of
K. Abbasi
(Nordmann, 1840)

Smitt, 1900
(De Filippi, 1863)


Members of this genus are characterised by an elongate body which is compressed posteriorly, a depressed head, a long snout, usually small scales on the occiput, short soft dorsal and anal fins both with 9-10 rays, the first dorsal fin with 6 spines, scales are ctenoid and number 23-40 in lateral series, anterior nostrils are tubular, the tongue is not notched, and teeth are simple.
There are about 48 species in Japan, Korea, Taiwan, China and the Amur River basin of eastern Asia. One species, presumably from the Amur River basin in the former Soviet Union, has been introduced to Turkmenistan and has reached Iranian waters.
Rhinogobius similis
Gill, 1859


Anzali Wetland, October 2011, courtesy K. Abbasi
Common names
gel-ye mahi, gavmahi-ye talabi (= pond or lake goby).
[lake goby]
Systematics
Rhinogobius similis was originally described from Shimoda, Japan
Rhinogobius similis lindbergi Berg, 1933 described from the Amur and Ussuri rivers, Russia is the subspecies of the Amur River basin according to Pinchuk (1978) but Eschmeyer (1998) records it as a synonym of the type subspecies. The Iranian specimens probably had their ultimate origin in the Amur River basin. Vasil'eva (2007) and Vasil'eva and Kuga (2008) consider that the Rhinogobius introduced to Central Asia are R. cheni (Nichols, 1931), a Chinese species of the Yangtze River, and this could be the species found in Iran. However, the large number of species in the genus, the poor state of knowledge on these fishes and the source presumably being the Amur River, leaves this identification as open.
Key characters
The only member of its genus in Iran, its key characters are those outlined in the genus.
Morphology
Dorsal fin spines 5-8, dorsal soft rays 7-10 after an initial unbranched ray, anal fin soft rays 6-10 after an initial unbranched ray, lateral series scales 28-36. Scales can be absent from the midline of the nape and the parietal and occiptal regions but this is variable (the nominal subspecies lindbergi lacks scales). Adult fish have the dorsal spines elongated (but not in lindbergi, not even in males). Scales in small Iranian specimens are a vertical oval, the posterior margin is fringed with ctenii, circuli in these small specimens are moderately devloped, radii radiate from the central point of the posterior margin to the anterior margin, the posterior margin has almost straight upper and lower margins coming to a central point so flank scales form a diamond pattern. The gut is a short s-shape. Gill rakers number 8-12 and are short and touch the adjacent one when appressed. Total vertebrae number 24-28.
Meristic values for Iranian specimens are:- first dorsal fin spines 6(3); second dorsal fin soft rays 9(3); pectoral fin rays 18(2) or 19(1); lateral series scales 31(3); total gill rakers 10(1) or 11(2); and total vertebrae 27(2).
Sexual dimorphism
Unknown.
Colour
Overall colour is a pale yellow or light brown with 6 brownish blotches on the flank and 6 blotches on the back, their position alternating. A dark band runs through the eye to the posterior edge of the mouth. The upper head is speckled brown. The belly is white. There is a dark spot between the first and second dorsal fin spines in the nominal lindbergi subspecies. There are 2 rows of spots on the first dorsal fin and 3-4 more distinctive rows on the second dorsal fin. The caudal fin has 2-5 rows of spots. The anal fin bears pigment distally on the membranes. The pectoral and pelvic fins are greyish. The pigmentation of the type subspecies is given by Pinchuk (1974) but may differ in detail from the nominal lindbergi subspecies so is not given here.
Young fish have 6 blotches along the mid-flank and membranes of fins are either strongly pigmented overall or bear up to 6 bars on the first dorsal fin, second dorsal fin and caudal fin with pectoral, pelvic and anal fins much lighter, almost immaculate. There is a small blotch between the first and second first dorsal fin rays near the fin base. The distal margin of the anal fin is yellowish-orange. The peritoneum is silvery but has a strong development of melanophores dorsally.
Size
Attains 100 mm.
Distribution
Aliev et al. (1988) refer to an exotic species of goby occurring in the Tedzhen River of Turkmenistan in the text of their work but not in the table of distributions. A goby is also reported by Shakirova and Sukhanova (1994) and Sal'nikov (1995) from the Karakum Canal and the Kopetdag Reservoir in Turkmenistan on the northern borders of Iran. Specimens have been collected from Sarakhs, Iran in the Tedzhen (= Hari) River (Coad and Abdoli, 2000a; Abdoli et al., 2000) and Abdoli (2000) records it from the Kashaf and Hari rivers in the Tedzhen River basin. K. Abbasi (pers comm., June 2012) records it from the Anzali Wetland.
Zoogeography
An exotic or alien species from eastern Asia, introduced to Turkmenistan from where it presumably entered Iranian waters. The genus Rhinogobius is in need of revision and the correct name for the taxon in Iran may change.
Habitat
The Iranian specimens were caught in the Hari River which had a depth of 35-120 cm , a width of 12-27 m, a turbidity reading of 15 cm, a water temperature of 12°C and a coarse gravel bed. It eastern Asia it is found in both rivers and lakes (hence the common name of lake goby as used in Japan). It is found on shallow sandy bottoms and on the upper surface of large stones in shallow water. Water temperatures reach 28°C (Kopylets and Dukravets, 1981).
Age and growth
Life span exceeds 4 years (Kopylets and Dukravets, 1981).
Food
Unknown.
Reproduction
Egg production reaches 1862 eggs in total (Kopylets and Dukravets, 1981).
Parasites and predators
Unknown for Iran.
Economic importance
Welcomme in Courtenay and Stauffer (1984) reports this species to be a pest when introduced.
Conservation
As an introduced species this fish requires no conservation.
Further work
The spread of this species in Iranian waters and its effects on native fauna should be documented.
Sources
Iranian material: Uncatalogued material, 5, 42.4-47.6 mm standard length, Khorasan, Hari River near Sarakhs (36º30'N, 61º10'E).
© Brian W. Coad (www.briancoad.com)